|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Границы применения гипотезы анализа признаков
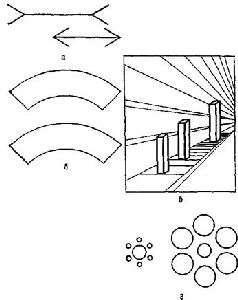
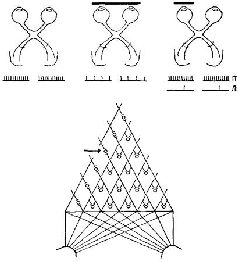
Почему же неправильно рассматривать перцептивные процессы как безобразный анализ признаков? По трем причинам. Во-первых, анализируемые признаки не являются какими-то особыми, отличными от других. Во-вторых, богатство феномена восприятия не объясняется теми детекторами признаков, которые открыты. И в-третьих, изменения, которые имеют место в системах входа в ходе развития организма, позволяют различать эффекты, возникающие в детекторах признаков, изученные с помощью микроэлектродов, и эффекты, продуцируемые в системе, анализирующей эти признаки, что было изучено в поведенческих опытах с различением сигналов. Рассмотрим прежде всего проблему различимости признаков. И. Рок (1970) со всей проницательностью сформулировал эту проблему следующим образом: «… Те, кто утверждал бы, что ответ нейронных детекторов может объяснить, почему вещи выглядят такими, какими они являются на самом деле, возможно, не знают тех фактов, касающихся восприятия, на которые гештальтисгы обращали особое внимание. Например, известно, что воспринимаемая нами величина, не просто зависит от величины образа на сетчатке, воспринимаемая форма не является непосредственной функцией формы соответствующего изображения, а воспринимаемая скорость — непосредственной функцией скорости перемещения изображения на сетчатке и так далее. Напротив, несмотря на значительные изменения находящегося в отдалении от нас стимула, возникающие при изменении позиции наблюдателя, предметный мир сохраняет удивительную константность. Эти факты нельзя отрицать, и их нельзя оставить без внимания на том основании, что вещи в этих изменяющихся условиях не выглядят константными, а мы знаем, что они константны. Против этой формулировки имеются достаточно веские возражения. Рассмотрим детально некоторые факты, касающиеся восприятия движения. Начнем с того, что существует константность положения, а именно-известно, что, когда мы перемещаемся или двигаем глазами, перемещается и образ всей картины на сетчатке, но сама картина не кажется движущейся. Случается также обратное, а именно неподвижный образ на сетчатке вызывает восприятие движения при движении наших глаз или головы, как это наблюдается при слежении глазами за движущимся объектом или при рассматривании последовательного образа во время движения тлаз. Известно, далее, стробоскопическое движение, когда сменяющиеся неподвижные изображения вызывают ощущение движения. Известен также феномен индуцированного движения предмета, изображение которого на сетчатке неподвижно. Этот феномен возникает вследствие перемещения окружающих объектов. Так, например, Луна кажется движущейся на фоне перемещающихся облаков. Можно принять гипотезу, что действительной сенсорной основой восприятия движения является стимуляция детекторов движения, но тогда необходимо предположить, что значимость такой стимуляции определяется другой информацией. Например, можно сказать, что движение вообще не воспринимается, если данным о смещении изображения на сетчатке противостоит информация о том, что оно возникает в результате собственного произвольного движения организма, как это предположил фон Хольст, — а до него Гельмгольц. Или можно считать, что в случаях индуцированного движения детекторы предоставляют решающую информацию о том, что что-то движется, но по неизвестным еще причинам движение приписывается неподвижному объекту. Откровенно говоря, я не считаю это достаточно убедительным. Это не объясняет, почему я вижу свой последовательный образ движущимся, когда я двигаю глазами даже в совсем темной комнате, где не видно неподвижных предметов, изображения которых могли бы перемещаться на сетчатке. В случае же стробоскопического движения, если наши глаза удерживаются в стационарном положении, то источником стимуляции детекторов движения мог бы быть только возникающий в этих условиях нистагм глаза. Но это не могло бы объяснить движения, которое возникает между двумя раздельно освещаемыми объектами. Эта гипотеза не может объяснить и индуцированного движения, потому что предмет, который вызывает ощущение движения, может двигаться так медленно, что его движение будет ниже порога, в то время как только оно и воспринимается. Далее, очевидно, что в данном случае основой восприятия движения является изменение локализации предмета, который кажется движущимся относительно местонахождения предмета, который вызывает это движение, а не стимуляция детекторов перемещения изображения. Рассмотрим теперь следующую проблему — проблему восприятия ориентации предмета. Является ли основой воспринимаемой ориентации линии ориентация ее изображения на сетчатке? Очевидно, нет, поскольку при повороте головы вертикальные и горизонтальные линии на стене продолжают оставаться для нас вертикальными и горизонтальными. Здесь мы «нова сталкиваемся с константностью, в данном случае с константностью восприятия направления, несмотря на изменения в ориентации изображений на сетчатке. Это происходит в значительной степени даже тогда, когда мы рассматриваем в темноте одну светящуюся линию, наклонив или откинув голову в другую сторону. Следовательно, обоснованно заключение, что, прежде чем сделать вывод о значении для восприятия ориентации данного сетчаточного образа, необходимо принять во внимание информацию о собственной позиции наблюдателя. Далее, как бы мы поняли открытие, сделанное Хьюбелом и Визелом, о существовании в зрительной коре детекторов ориентации? Очевидно, что они не могут быть знаками ориентации объектов в окружающей среде. Можно согласиться, что они являются знаками, указывающими на ориентацию объектов относительно организма, другими словами, детекторами эгоцентрической ориентации, и что они создают условия, необходимые для того, чтобы организм мог определить ориентацию объектов в окружающей среде, когда он дополнительно получает информацию и об ориентации самого наблюдателя. Например, можно сказать, что, когда изображение линии на сетчатке будет иметь вертикальное положение, оно всегда будет казаться эгоцентрически вертикальным, то есть параллельным продольной оси головы, и линия будет восприниматься вертикальной в поле зрения наблюдателя, если он занимает вертикальное положение, и горизонтальной, если наблюдатель повернут на 90 градусов. Хотя такое объяснение кажется мне правдоподобным, оно вызывает серьезные трудности из-за существования некоторых дополнительных фактов. Если наблюдатель сохраняет вертикальное положение, то вертикальная линия, воспринимаемая им на фоне изображения комнаты, при наклоне последнего будет выглядеть наклоненной в направлении, противоположном наклону изображения комнаты. Таким образом, в данном случае вертикальное изображение на сетчатке не будет более соответствовать вертикально расположенному предмету, несмотря на то, что наблюдатель находится в вертикальном положении. Более того, линия не будет уже казаться эгоцентрически вертикальной линией. К этим фактам можно было бы добавить данные о том, что наблюдатели будут адаптироваться к повернутым с помощью призматических стекол изображениям, так что вертикальное изображение на сетчатке не будет больше ни знаком вертикального объекта, когда наблюдатель находится в вертикальном положении, ни фактически знаком того, что объект ориентирован в вертикальном направлении относительно наблюдателя. Имеют ли какое-либо отношение детекторы ориентации в коре к восприятию формы? Можно предположить, что восприятие формы сводится к восприятию ориентации линий, из которых строится форма. Таким образом, предмет воспринимается как квадрат тогда, когда одновременно стимулируются два детектора вертикальных и два детектора горизонтальных линий. Хорошо известно, что фигуры выглядят весьма по-разному, когда меняется их ориентация. На первый взгляд сам этот факт может казаться доказательством того, что детекторам ориентации принадлежит существенная роль в восприятии формы, потому что в этом процессе принимают; участие совершенно различные волокна. Можно сказать, что это является основой для понимания явления изменения внешнего вида предмета. Однако в действительности, вообще говоря, объяснением изменения внешнего вида предмета служит не изменение ориентации изображения на сетчатке. В проведенных мною и моими сотрудниками экспериментах неоднократно было показано, что, если положение фигуры в отношении ее окружения не меняется, но она рассматривается наблюдателем в наклонной позе, фигура не выглядит измененной, или, выражаясь более точно, распознавание не ухудшается. Могло ли бы это быть, если за это явление были бы ответственны совершенно различные детекторы? Напротив, если фигура наклонена так же, как и наблюдатель, и ориентация изображения на сетчатке, таким образом, совсем не изменилась, фигура тем не менее выглядит совершенно иной и иоэтому часто не узнается. Можно провести простой опыт: создайте последовательный образ квадрата, держа голову в вертикальном положении, а затем, закрыв глаза и повернув голову на 45 градусов, посмотрите на него, он будет подобен ромбу. Могло ли бы это быть, если за это явленяе были бы ответственны одни и те же детекторы? Однако эти факты приобретают смысл, если считать, что решающим — для ориентации является восприятие определенной части фигуры как верх, а другой — как низ и так далее. Если мы будем правильно оценивать одну и ту же часть фигуры в качестве верха, как мы это обычно делаем, приняв наклонное положение, то мы увидим, что форма предмета не меняется. Однако, если мы за верх фигуры принимаем другую ее часть, как это бывает при наклоне фигуры относительно окружения, то мы увидим, что ре форма очень изменилась. Этот процесс, по-видимому, не имеет ничего общего со специфическими детекторами ориентации. Далее, роль этих детекторов, возможно, состоит в том, что они служат для различения ориентации одного образа по отношению к другому. В прошлом мы ошибочно считали, что такое различение осуществляется благодаря различной пространственной ориентации проекционных кортикальных «образов». Иными словами, нейронные детекторы, вероятно, являются механизмами, посредством которых воспринимается определенная информация о стимуле, в данном случае информация о том, что сохраняется или изменяется пространственная ориентация. Конечно, такая информация необходима для восприятия в качестве исходной, но ответ волокон детекторов никоим образом нельзя рассматривать как нейронные корреляты единиц восприятия» (1970, р. 2—5). В остальной части статьи Рока анализируется еще ряд интересных экспериментов, которые трудно объяснить с позиции детекторов признаков (см. также Pис. VII—10, VII—11).
Второй недостаток точки зрения, рассматривающей восприятие как безобразный процесс выделения признаков, состоит в том, что по сравнению с богатством явлений сознания процесс распознавания структуры, осуществляемый методом классификации, довольно беден деталями, которыми он оперирует. Механизм распознавания зрительной структуры, который осуществляется путем избирательного выделения признаков, может приводить к ошибкам. Каждый из нас ждал кого-нибудь, кто опаздывал на свидание, назначенное в многолюдном месте, и ошибочно принимал посторонних за ожидаемое лицо, так как они имели сходные с ним черты. Мы узнаем А, Л и а как букву А; мы хорошо знаем о различиях почерков. Восприятие, по-видимому, обеспечивает в одно и то же время не только богатство нашего опыта и «сильное предпочтение сохранять постоянную организацию», но также и тенденцию к «максимальному упрощению», как показали опыты гештальтпсихологов. Имеются данные, свидетельствующие о том, что богатство восприятия связано со сложностью конфигураций окружающей среды, которые существуют независимо от организма и возбуждают его рецепторные поверхности (см. Gibson, 1966), последние приводят в действие некоторый механизм мозга с соответствующей емкостью, позволяющей вместить это богатство. Данные, представленные Роком и другими, показывают, что только механизм выделения признаков обусловливает тенденцию к упрощению, создает схему отношений. Однако даже здесь начинает играть роль другой механизм — более сложный «познавательный» процесс.
Третий недостаток объяснения фактов восприятия с позиции выделения признаков выступает в экспериментах, в которых новорожденные в течение различных периодов времени подвергались сенсорной депривации. Как уже говорилось в главах VI и VIII, младенцы в возрасте нескольких недель обнаруживают способность к восприятию константности величины и формы. У животных способность к различению развивается также рано; при изучении мозга детенышей животных были обнаружены детекторы признаков, которые уже достигли своего полного развития. Однако при ограничении потока сенсорных воздействий эта простая корреляция между поведением и функциями мозга нарушается. Так, когда кошки и крысы содержатся в полной темноте, их способность различать яркость и контур остается неизмененной, но способность различать более сложные зрительные структуры значительно ухудшается. Хотя ориентация рецептивных полей кортикальных нейронов у таких животных еще различима, она имеет менее острую настройку, и утомление клеток развивается быстрее, чем у животных, содержащихся в нормальных условиях. Но когда сенсорная депривация ограничена депривацией одного глаза в течение определенного времени, как зрительная ориентация, так и способность к различению оказываются резко нарушенными, если животное пользуется только одним этим глазом. Это связано с тем, что стимуляция глаза, подвергнутого депривации, может привести в активное состояние лишь небольшое число нейронов, которые вряд ли могут сколько-нибудь избирательно отвечать на определенным образом ориентированную линию или движение. Эта реакция возникает, даже несмотря на то, что в коре присутствует обычный для нее комплект клеток с их нормальной реактивностью, как это было обнаружено при стимуляции не подвергавшегося депривации глаза (см. Рис. VII—12). Таким образом, когда имеет место нарушение в привычном способе возбуждения, происходит какое-то подавление реактивности и это подавление переходит в нарушение функции исключительно в результате неупотребления нервных структур (как у животного, подвергшегося полной сенсорной депривации). Если монокулярное подавление реактивности коры объясняется некоторым изменением в связях, то ответственным за этот эффект подавления должен быть какой-то процесс, отличный от простого выделения признаков. Л. Ганц в недавнем обзоре (1971) этих и других сходных данных доказал это. Основываясь на результатах экспериментов с полной депривацией животных, он различает два типа заданий. Ориентация по яркости и контуру требует только, чтобы организм отвечал на языке реакций, свойственных большинству его детекторов признаков. Различение более сложных структур, например двух треугольников, один из которых перевернут, не может осуществляться на этой основе. Мы нуждаемся в каком-то дополнительном, более гибком мозговом механизме, который Ганц называет «избирательным вниманием». Этот механизм активно подавляет доминирующую реакцию детекторов признаков, реакцию, которая, в сущности, не имеет отношения к адекватному решению задачи на различение. Следовательно, мы снова сталкиваемся с необходимостью иметь дополнительный механизм, помимо выделения признаков. Поэтому мы снова приходим к мысли о той, что, возможно, системы входа организованы таким образом, что нейронные сигналы координируются с каким-то психологическим процессом формирования образов. Проблема состоит в том, чтобы определить нейро-логическую природу этой организации. На основе рассмотренных в начале этой главы данных становится очевидно, что эта организация не может быть просто мозаикой точек, которые генерируются в рецепторах и без изменения проводятся до коры, где они формируют образ, подобно фотографии. Столь же очевидно, что следует принять во внимание наличие нейронов, чувствительных к линиям, и других детекторов признаков, которые создают в мозгу нейронную основу для их анализа. Резюме Почти полное удаление мозговой ткани независимо от ее локализации в пределах систем входа оказывает лишь незначительное влияние на процесс распознавания. Следовательно, логика работы нейронов, свойственная любому конкретному восприятию, распространяется, по-видимому, на всю систему. Были получены также прямые данные, касающиеся распределения памяти в мозгу. Логику этой памяти создают как встроенные детекторы элементарных признаков (таких, как линии и углы), так и анализаторы для более сложных преобразований особых признаков (таких, как волновые формы), которые перестраиваются в ходе жизненного опыта организма. Главная » Карл Прибрам. Языки мозга. Экспериментальные парадоксы и принципы нейропсихологии
Поиск по сайту: |