|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
ВЧЕННЯ І.П. ПАВЛОВА ПРО ВИЩУ НЕРВОВУ ДІЯЛЬНІСТЬСтр 1 из 3Следующая ⇒
Зміст [сховати] · 1 Роль у роботі ЦНС o 1.1 Ритми ЕЕГ o 1.2 Центральні генератори впорядкованої активності o 1.3 Пам'ять o 1.4 Сон o 1.5 Увага · 2 Патологія o 2.1 Пов'язані захворювання § 2.1.1 Епілепсія § 2.1.2 Хвороба Паркінсона § 2.1.3 Когнітивні розлади o 2.2 Методи боротьби § 2.2.1 Фармацевтичні § 2.2.2 Хірургічні § 2.2.3 Метод глибокої стимуляції мозку · 3 Математичне моделювання o 3.1 Мотивація o 3.2 Моделі біологічного нейрона § 3.2.1 Модель Курамото § 3.2.2 Спайкові моделі o 3.3 Види синхронізації осциляторів o 3.4 Глобальна функція стабільності синхронізованого стану · 4 Джерела Роль у роботі ЦНС[ред. • ред. код] Ритми ЕЕГ[ред. • ред. код]
Приклад спектру ЕЕГ (чорний графік). Загальна потужність кожного ритму дорівнює площі відповідного сірого прямокутника. Зображення адаптовано з van Albada SJ and Robinson PA, 2013[3]. Електроенцефалографія (ЕЕГ) — один з ключових неінвазивних методів дослідження головного мозку, який характеризується високою часовою роздільною здатністю, але досить низькою просторовою роздільною здатністю, особливо у порівнянні з МРТ або інвазивними електродними методами. Через низьку просторову роздільну здатність (зазвичай не більше як 100 каналів), отриманий сигнал є результатом суперпозиції активності великих популяцій нейронів, а значить є зручним методом для знаходження та дослідження синхронізованих колективних режимів у мережах головного мозку. Синхронізація достатньо великої популяції нейронів зазвичай продукує глобальні коливання, які й записуються електродами електроенцефалографа. У більшості випадків, сильна глобальна синхронізація великих популяцій (синхронізація типу I) притаманна стану бездіяльності або стану патологічному, адже динаміка повністю синхронізованої мережі не достатньо складна для ефективної обробки інформації. Під час нормальної ж роботи (окрім стану глибокого сну), невеликі локально-синхронізовані підмережі генерують коливання різних частот (синхронізація типу II), при цьому глобальна синхронізація зникає[4]. Таким чином, спектр ЕЕГ сигналу містить декілька важливих складових, які прийнято класифікувати за частотою, призначаючи кожному діапазонові грецьку літеру. У таблиці нижче наведено короткий опис кожного з ритмів, відповідно до традиційної системи класифікації, а також їх відповідні функції. У наступних секціях більш детально розглянуто властивості та шляхи утворення деяких ритмів.
Окрім вищезгаданих канонічних ритмів, сигнал ЕЕГ може також містити високочастотні компоненти, з частотою понад 400 Гц. У більшості випадків, такі компоненти мають дуже низьку енергію та часто нехтуються або сприймаються як шум. Однак нещодавні дослідження показали, що високочастотні коливання в сигналі ЕЕГ, які можна спостерігати в соматосенсорній корі головного мозку у відповідь на стимуляцію периферійного нерву, можуть бути результатом дуже точної синхронізації нейронних сплесків у відповідних мережах[9]. В експериментах з одночасним отриманням ЕЕГ та зовнішньоклітинного сигналу, було показано, що форма високочастотних компонентів сигналу ЕЕГ містить інформацію про часову структуру сплеску спайків індивідуальних нейронів. Таким чином, високочастотні ритми ЕЕГ можуть пролити світло на динаміку клітинного рівня, без втручання в біологічні тканини[9]. Центральні генератори впорядкованої активності[ред. • ред. код] Синхронізована активність нейронів є базою для генерації періодичних моторних команд для ритмічних рухів. Ритмічні команди у свою чергу виробляються групою зв'язаних нейронів що формують мережу, яка називається центральний генератор впорядкованої активності (ЦГВА). При активації, такі нейронні мережі можуть генерувати складні ритмічні моторні команди, навіть за відсутності сигналів зворотнього зв'язку, що несуть інформацію про конкретні часові інтервали. Ходіння, плавання або диханняє прикладами роботи ЦГВА[10]. ЦВГА найбільш досліджені у нижчих тварин, таких як міногові, проте також існують свідчення про наявність ЦВГА у людей[11]. Пам'ять[ред. • ред. код] Докладніше: Пам'ять Фазова синхронізація (ФС) коливань між різними зонами мозку має ряд важливих функцій, які є критичними для ефективної роботи пам'яті. Важливою функцією є покращення передачі інформації між віддаленими ділянками мозку під час ФС[1]. Оптимальна передача інформації може відбуватися завдяки ФС наступним чином: високий рівень локального потенціалу зазвичай свідчить про високий рівень активності в локальній популяції в даний момент часу. Одночасно, чутливість нейронів цієї популяції також підвищена, бо мембрани нейронів у середньому більш деполяризовані ніж коли локальний потенціал має низькі значення. Таким чином, якщо дві віддалені популяції нейронів мають синхронізовані колективні коливання, то момент підвищеної активності одної популяції збігатиметься з високою збудливістю іншої, забезпечуючи високу імовірність передачі інформації[12]. ФС також може сприяти підсиленню синаптичної пластичності. Синаптична пластичність що залежить від часу імпульсу (STDP) спричиняє довготривалу потенціаціюсинапсів, якщо приймаючий нейрон спрацьовує не більше як 10-20 мс після вхідного імпульсу. ФС може забезпечити достатньо вузькі інтервали спрацьовування нейронів між двома групами і таким чином збільшити або зменшити силу зв'язку між ними[1]. За поширеною системою класифікації, існують два основних види пам'яті: · Короткочасна пам'ять · Довготривала пам'ять У роботі короткочасної пам'яті, ФС спостерігається між префронтальною корою та скроневою долею мозку (на частоті θ-ритму) під час читання, запису та утримування короткочасних спогадів, а також між фронтальними та паріетальними зонами кори (на γ та β-частотах) під час утримування спогадів. Запис та читання візуальних об'єктів із декларативної пам'яті (тип довготривалої пам'яті) супроводжуються фазовою синхронізацією коливань між антериальними та постериальними зонами мозку на θ, γ, δ та β-частотах. Інші види синхронізації також беруть участь у репрезентації об'єктів у пам'яті[1]. Результати експериментів з одночасним записом електричної активності в гіпокампі та латеральній мигдалеподібній залозі свідчать про підсилення синхронізації на θ-частотах між цим ділянками після формації та під час дії умовного рефлексу страху[13]. Сон[ред. • ред. код] Докладніше: Сон
Дельта-ритм (у червоній рамці) під час глибокої фази сну. Під час глибокої фази сну, а також при деяких видах анестезії, на енцефалограмах зазвичай спостерігаються коливання низької частоти (0.3 — 1 Гц) та дельта коливання. Такі ритми часто асоціюються з відсутністю свідомості, а також часто використовуються як індикатор наявності свідомості у пацієнта під наркозом. На клітинному рівні, осциляції виглядають як періодична зміна станів активного спрацьовування нейронів та тиші, кожен з яких триває до одної секунди[14]. Одним із можливих механізмів генерації повільних коливань (до 1 Гц) є спільна робота відповідних мереж таламусу та неокортексу, наступним чином: · активність нейронів в корі ініціює внутрішні осциляції в таламусі · коливання в таламусі посилюють синхронізацію гальмівних інтернейронів · активація гальмівних інтернейронів пригнічує активність нейронів кортексу і настає період тиші · цикл повторюється Інші механізми утворення низькочастотних коливань добре освітлено у роботі, на приклад, М. Тимофеєва[14]. При нормальних умовах, глибокий сон відіграє важливу роль у процесах консолідації пам'яті. Відповідно до популярної теорії, повільні колективні коливання регулюють ре-активацію та транспорт спогадів з гіпокампу докортексу. Під час фази швидкого сну (підвищена тета-активність та локальна синаптична пластичність), ре-активовані спогади у кортексі закріплюються на синаптичному рівні, тобто відкладаються у довготривалу пам'ять[15] [16]. Увага[ред. • ред. код] Докладніше: Увага Увага — спрямованість психічної діяльності людини або тварини та її зосередженість у певний момент на об'єкти або явища, які мають певне значення при одночасному абстрагуванні від інших, в результаті чого вони відображаються повніше, чіткіше, глибше, ніж інші[17]. Під час концентрації уваги на конкретному об'єкті або його властивості, підвищується швидкість обробки інформації, зменшується час реакції, підвищується рівень точності, чутливості до невеликих змін у стимулі та сприйняття контрасту. По перше, увага підсилює активність нейронів що репрезентують об'єкт уваги або певну характеристику (колір, орієнтація, тощо). Так, на приклад, нейрони зорової кори,рецептивне поле яких містить об'єкт уваги, сильніше синхронізовані з локальним потенціалом (на γ-частоті), ніж нейрони що реагують на інші об'єкти поза уваги. Такий спосіб селективного підсилення синхронізації імовірно є альтернативою підвищення частоти спрацьовування нейронів (вона зазвичай не змінюється в залежності від спрямування уваги)[18]. По друге, увага також регулює інформаційне сполучення між віддаленими частинами мозку, підсилюючи зв'язок між групами нейронів що несуть важливу для тварини інформацію. Багато досліджень приходять до виводу що таке підсилення зв'язку відбувається за рахунок селективної синхронізації між зонами кори[19]. Патологія[ред. • ред. код] Під час нормальної роботи мозку, локальна та міжзонна синхронізація нейронної активності грають важливу роль у таких ключових процесах, як увага, пам'ять, моторика тощо. Однак важливим є також і певний баланс синхронізації та десинхронізації[20]. Надмірна або недостатня синхронізація може бути причиною багатьох патологічних процесів, в числі яких епілепсія, тремор, шизофренія, деменція, та деякі інші. Таким чином, важливим напрямком розвитку нейронауки є пошук ефективних шляхів знешкодження патологічної синхронізації, та розуміння принципів її утворення. Пов'язані захворювання[ред. • ред. код] Епілепсія[ред. • ред. код] Докладніше: Епілепсія
Електроенцефалограма, характерна для епілептичного припадку. Епілепсія характеризується раптовим початком синхронізації у відносно великих нейронних мережах, внаслідок чого стає неможливою ефективна обробка інформації, що в свою чергу перешкоджає нормальній роботі мозку. Епілептичні припадки можуть супроводжуватись короткочасною втратою свідомості або більш складними сенсорними та моторними проявами[20]. Найбільш проблематичним наслідком епілепсії є непередбачуваність припадків, внаслідок чого людям, що страждають епілепсією, може бути на законодавчому рівні заборонено керувати автотранспортом та виконувати деякі інші заняття, що вимагають постійної уваги. Точні нейронні механізми епілептичної синхронізації наразі досконало не відомі. Проте застосування теорії динамічних систем до аналізу та моделювання сигналу що генерується такими мережами дає певні результати у напрямку розуміння феномену. Будь-яка нейронна мережа може бути описана як (нелінійна) динамічна система з одним або декількома атракторами. (Атрактор — це множина точок у фазовому просторі системи, до якої збігаються траєкторії системи. Іншими словами, це тип поведінки, до якого система повертається після невеликого відхилення в результаті зовнішнього втручання. Найпростішим прикладом атрактора є граничний цикл. На такому атракторі система починає описувати періодичні коливання). Згідно з однією з теорій, перехід від нормальної до епілептичної активності мережі може бути двох видів[21]: · В результаті поступової деформації атрактора нормальної активності в атрактор активності епілептичної (через відповідну біфуркацію), внаслідок зміни параметрів системи (наприклад, зовнішньоклітинної концентрації іонівкальцію або магнійю), або зовнішнього стимулу. Така модель відповідає типам припадків, коли зміна стану протікає поступово, у декілька фаз. · Внаслідок різкого «перескакування» системи з одного нормального атрактора на атрактор синхронізованої активності, що було спричинено випадковою флуктуацією або зовнішнім стимулом (наприклад, спалахами світла). На відміну від першого випадку, тут система одночасно має два або декілька атракторів, включаючи нормальні і епілептичні (таку систему називаютьмультистабільною). В цьому випадку, в мозку здорової людини такі атрактори можуть бути розташовані далі один від одного, що унеможливлює випадковий перехід в стан епілептичного припадку). В залежності від типу епілепсії, можуть бути застосовані різні підходи до моделювання динамічних процесів, що призводять до масової синхронізації нейронів[21] [22] [14]. Хвороба Паркінсона[ред. • ред. код] Докладніше: Хвороба Паркінсона
Сильний тремор лівої руки у пацієнта з хворобою Паркінсона. Хвороба паркінсона — це повільно прогресуюче рухове порушення, яке супроводжується порушенням мови, скутістю, підвищенням тонусу м'язів, тремором та ригідністю м'язів, сповільненням рухів. Тремор, що характеризується ритмічними неконтрольованими рухами кінцівок, є дуже проблематичним симптомом, так як, при певних умовах, унеможливлює виконання хворим багатьох повсякденних дій, таких як керування авто, утримування відкритої ємкості з рідиною, рукописання тощо. Патологічний процес при цьому захворюванні характеризується деградацією дофамінергічних нейронів чорної речовини, що тягне за собою посилення активності та синхронізації нейронів в субталамічних ядрах (частина базальних ганглій) і призводить до надмірного гальмування таламо-кортикального зв'язку. Наразі, точні механізми генерації тремору у пацієнтів з хворобою Паркінсона невідомі. Однак все більше даних кореляційного характеру свідчать про зв'язок між абнормально синхронізованою активністю в певних частинах мозку та м'язовим тремором[23][24]. Так, відповідно до провідної гіпотези, тремор генерується групою нейронів у базальних ядрах, які працюють як центральні осцилятори та продукують періодичні імпульси певним групам м'язів. При чому важливим є як загальний рівень активності центральних осциляторів, так і рівень їх фазової синхронізації[23]. Когнітивні розлади[ред. • ред. код] Шизофренія — це когнітивний розлад, головними симптомами якого є галюцинації, сильно спотворене сприйняття реальності, та загальна дезорганізація мислення та дій. Як і у випадку з багатьма складними ментальними захворюваннями, патофізіологія шизофренії досі залишається до кінця не відомою[25]. Як відомо, осциляторна активність мереж головного мозку пов'язана з такими важливими процесами як селективність до зовнішніх стимулів за допомогою уваги, пам'ять та свідомість. Так як симптоми шизофренії пов'язані саме з розладами вищезгаданих процесів, є підстави вважати, що абнормально низька β та γ-синхронізація, що спостерігається у хворих, є важливим фактором у патофізіології захворювання. Сучасні емпіричні дані свідчать про нелокальний та системний характер дисфункції головного мозку, що виникає при шизофренії, та може включати у себе розлад в динаміці нейронних мереж у префронтальних та тім'яних зонах, а також погіршення зв'язків між віддаленими зонами[25][26]. Аутизм, захворювання що пригнічує соціальну та комунікативну здібності пацієнтів, також було пов'язано зі зменшеною осиляторною активністю на γ-частотах. Хворі на аутизм страждають звуженим фокусом уваги та не здібні достатньо добре інтегрувати фрагменти інформації у єдине ціле, що може бути наслідком недостатньо ефективної передачі інформації між зонами мозку[25][26]. Методи боротьби[ред. • ред. код] Фармацевтичні[ред. • ред. код] Одним з поширених типів фармацевтичного пригнічення надмірної синхронізації, є використання різноманітних блокаторів іонних каналів, внаслідок чого зменшується збудливість цільових нейронів або ж пригнічуються синаптичні зв'язки. Наприклад, збільшення зовнішньоклітинної концентрації Mg2+ може припиняти синхронізовану коливальну активність в моделях епілепсії. З одного боку, це відбувається завдяки здатності іонів магнію блокувати NMDA-рецептори в синаптичних зв'язках між нейронами. З іншого боку, магній, та деякі інші 2 та 3-валентні іони (наприклад, Ca2+) можуть впливати на локальних заряд навколо натрієвих та калієвихпотенціалчутливих каналів, знижуючи активаційний поріг та деякі динамічні характеристики нервової клітини[27] [28]. Фармацевтичних засоби також можуть діяти через зв'язування та деактивацію певних нейротрансмітерів. Хірургічні[ред. • ред. код] Майже третина пацієнтів з епілепсією не показують позитивної динаміки в результаті лікування антиепілептичними медикаментами[29]. Однак в деяких випадках (близько 7-8%), вдається достатньо точно локалізувати джерело абнормальної епілептичної синхронізації в мозку, яке видаляється за допомогою оперативного втручання. Сучасні хірургічні методи також дозволяють проведення операцій з видалення надмірно синхронізованих популяцій в глибокій сіра речовині (бліда куля, субталамічні ядра), для усунення симптомів багатьох моторних захворювань, включаючи хворобу Паркінсона[30]. Метод глибокої стимуляції мозку[ред. • ред. код]
Електроди DBS на рентгеноскопічномузнімку черепа людини. Розвиток комп'ютерних технологій відкрив нову еру у лікуванні пацієнтів, які страждають на хворобу Паркінсона. Все частіше застосовується новий метод глибокої стимуляції мозку (Deep Brain Stimulation). Метод DBS полягає у використанні імпульсів електричного стимулятора, вбудованого поверхово в організм пацієнта. Через введений в мозок спеціальний електрод, імпульси подаються в базальні ганглії, зокрема, у субталамічні ядка, та виконують високочастотну стимуляцію груп нейронів, що призводить до помітного поліпшення у перебігу хвороби. Внаслідок суттєвого зменшення симптомів хвороби, пацієнти можуть повернутися до нормальної повсякденної діяльності та значно зменшити обсяг вживання ліків. Крім хвороби Паркінсона, показанням до операції є вроджені дистонії і есенціальний тремор[2]. Клінічні випробовування також тривають для застосування DBS у лікуванні депресії. Не зважаючи на великий успіх DBS у лікуванні багатьох хвороб, технологія поки що знаходиться на ранньому етапі розвитку, та повна відмова від хірургічних втручань на користь DBS є темою дебатів[30]. Математичне моделювання[ред. • ред. код] Мотивація[ред. • ред. код] Методи математичного моделювання фізичних та хімічних процесів знайшли широке застосування в біофізиці та нейронауці. Починаючи від моделювання стохастичної динаміки окремих іонних каналів і закінчуючи мережами зі складною неоднорідною топологією та мільйонами нейронів, моделі допомагають науковцям зрозуміти механізми функціонування нервової системи на різних рівнях абстракції, а значить і знайти нові шляхи лікування відповідних захворювань, та розробляти більш розумні алгоритми та обчислювальні машини. Головним призначенням математичної моделі є усунення неважливих факторів з якогось процесу або спостереження, та кристалізація саме того мінімального набору механізмів, який є достатнім для пояснення даних, в конкретному контексті. Правильно побудована модель дозволяє тестувати цікаві гіпотези, та робити важливі передбачення щодо роботи тої чи іншої системи. В контексті синхронізації, моделювання дозволяє застосовувати аналітичні та чисельні методи для оцінки динамічних режимів мережі. Наприклад, біфуркаційний аналіз детальної моделі нейронної мережі може допомогти визначити максимально допустимі значення фізіологічних параметрів, що не призведуть до переходу мережі у патологічно синхронізований режим. Моделі біологічного нейрона[ред. • ред. код] Біологічний нейрон можна моделювати на різних рівнях абстракції, але суть багатьох моделей полягає у виділенні квазіперіодичних властивостей його динаміки, тобто зображення нейрона як осцилятора певного рівня складності. Модель Курамото[ред. • ред. код]
Чисельне рішення системи з'єднанихосциляторів Курамото показує перехід до синхронізованого режиму. Модель з'єднаних осциляторів Курамото[31] — одна з найбільш абстрактних моделей призначених для дослідження синхронізації та коливань в нейронних мережах (осцилятори мають лише одну степінь свободи — фазу коливань, ігноруючи амплітуду)[32]. Взаємодії між такими осциляторами описуються простою алгебраїчною формою (такою як синус), та колективно генерують певну картину глобальної активності. Модель Курамото та її розширення (такі як наприклад додавання можливості формувати зв'язки різної сили) широко використовується для вивчення коливальних процесів у мозку[33]. Зокрема, її зручно використовувати для вивчення механізмів початку синхронізації в групі пов'язаних нейронів. У найпростішому вигляді, фаза (
де Завдяки простоті визначення, можна відносно швидко симулювати динаміку великих популяцій таких нейронів, а також знаходити аналітичні рішення в певних випадках. Спайкові моделі[ред. • ред. код] Спайкові моделі нейронів використовуються у випадках, коли не можна обмежитись лише фазовою або частотною абстракцією нейрона, тобто коли необхідно враховувати амплітуду коливань, та/або інші, більш детальні аспекти роботи біологічного нейрона. Характерною властивістю спайкових моделей є здатність генерувати активність, наближену до потенціалу дії (спайку). Такі мережі є дуже корисними, коли необхідно зробити передбачення щодо значень певних біологічних параметрів, бо в їх математичне визначення часто включені аналоги біологічних параметрів (наприклад, динаміка іонних каналів або концентрація певних речовин у зовнішньоклітинному середовищі). Однак головним недоліком таких моделей є складність та нелінійність їх математичного визначення, що обмежує можливості аналітичних розв'язків та вимагає суттєвих затрат комп'ютерного часу для рішень чисельних. Зазвичай один такий нейрон визначається системою нелінійних диференційних рівнянь. В обчислювальній нейронауці найбільш популярними є наступні моделі: · експоненційна модель «інтегрувати і спрацювати» (AdEx)[34]. Є найпростішою моделлю що здатна продукувати всі основні динамічні режими біологічного нейрону, такі як одноразове спрацьовування, регулярне спрацьовування у відповідь на стимуляцію постійним струмом, пачкова активність тощо. Широко використовується у дослідженнях синхронізації у великих гетерогенних популяціях нейронів. Описується системою з двох диференційних рівнянь. · Модель Хіндмарша-Роуза (HR) — одна з найпростіших неперервних моделей, що здатні генерувати пачкову, а також хаотичну активності. Описується системою з трьох диференційних рівнянь[35]. · Модель Годжкіна-Гакслі (HH) та похідні — сімейство детальних біологічно-реалістичних моделей нейронів, що включають у себе кінетику натрієвих, калієвих, кальцієвих та інших іонних каналів. Різні модифікації здатні дуже точно імітувати форму потенціалу дії справжніх нейронів. Система зазвичай описується щонайменш чотирма нелінійними диференційними рівняннями. Не зважаючи на складність математичного визначення, HH-подібні моделі успішно використовуються для досліджень ефектів синхронізації у великих популяціях нейронів[36]. Види синхронізації осциляторів[ред. • ред. код] Існує велика кількість різних визначень явища синхронізації, які можуть бути застосовані в тому чи іншому контексті. Відзначимо деякі основні види синхронізації · Повна синхронізація — найпростіший, з математичної точки зору, тип синхронізації, яких характеризується повною збіжністю відповідних змінних всіх нейронів у мережі. Наприклад, якщо i-тий нейрон описується вектором змінних · Фазова синхронізація накладає певні обмеження на фазові відношення між осциляторами, ігноруючи відношення амплітуд. Якщо у системи можна визначити поняття фази, то фазова синхронізація мережі таких осциляторів означає, що різниця фаз між елементами мережі повинна або взагалі не змінюватись з часом, або знаходитись в певних скінченних границях[37]. Такий підхід дозволяє досліджувати синхронізацію мереж з осциляторами, параметри яких можуть відрізнятися. · Узагальнена синхронізація. Як фазова, тільки відношення між фазами повинні описуватися певною функцією. · Пачкова синхронізація — синхронізація нейронів під час пачкової активності. Така синхронізація не вимагає точного співпадіння моментів спайків нейрноів, але вимагає синхронізованого спрацьовування пачок спайків(ритмічних, обмежених у часі сплесків активності з генерацією декількох потенціалів дії)[38]. Повна синхронізація Фазова синхронізація (з часовою затримкою) Пачкова синхронізація Типи синхронізації моделей нейронів (на прикладі двох) Глобальна функція стабільності синхронізованого стану[ред. • ред. код]
Глобальна функція стабільності синхронізованого стану (MSF) для мережі лінійно з'єднаних нейронів Хіндмарша-Роуза (HR). Зеленим кольором позначені ділянки функції, де повна синхронізація є стабільною (від'ємна максимальна експонента Ляпунова). Система може синхронізуватися, якщо всі власні числа матриці зв'язків мережі попадають у діапазон над зеленими ділянками.
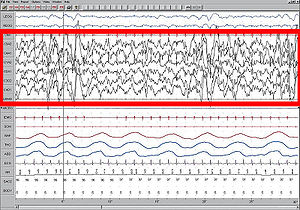
Повна синхронізація одномірної мережі (колонки) з 50 нейронів Хіндмарша-Роуза (HR). Сила синаптичного зв'язку експоненційно спадає з відстанню. Власні числа матриці зв'язків знаходяться в діапазоні [0.48, 0.5], тобто синхронізація стабільна (див. графік MSF вище). Верхня панель: час спрацьовування кожного з нейронів (сірі точки). Середня панель: Потенціал мембрани п'яти випадково обраних нейронів (у кожного свій колір). Нижня панель: апроксимація локального потенціалу популяції (середнє значення НЧ-відфільтрованих потенціалів мембрани). Простота математичного визначення стану повної синхронізації робить можливим застосування цікавих аналітичних методів для дослідження цього виду синхронізації. Глобальна функція стабільності синхронізованого стану (Master Stability Function, MSF), на приклад, дозволяє визначати стабільність стану повної синхронізації для мережі ідентичних нейронів[39]. Іншими словами, розрахувавши MSF для мережі потрібних моделей нейронів, можна з точністю сказати чи існують такі топології з'єднання нейронів, при яких дана мережа буде здатна синхронізуватися (тобто буде мати стабільний стан повної синхронізації). Краса методу полягає у тому, що MSF треба розраховувати лише один раз для кожного конкретного типу моделі та набору параметрів, а з результату можна робити висновки про стабільність синхронізованого стану для будь-якої кількості нейронів та будь-якої топології (при дотриманні умови однакової сумарної сили вхідного сигналу для нейронів). Основний процес аналізу синхронізації системи за допомогою MSF можна розбити на кілька кроків. Розглянемо мережу ідентичних зв'язаних нейронів, кожен з яких описується певною системою диф. рівнянь (наприклад, Модель Ходжкіна-Хакслі). Тоді мережу з N нейронів можна записати у такому вигляді:
де Отож, загальна розмірність фазового простору всієї мережі дорівнюватиме
де Опустивши деталі, маємо наступний алгоритм: 1. Розрахувати максимальну експоненту Ляпунова ( 2. Побудувати графік залежності 3. Знайти власні числа матриці зв'язків 4. Якщо всі власні числа матриці зв'язків (за винятком того, яке дорівнює рядковій сумі матриці, див.[39]) попадають у діапазон Приклад графіку MSF для HR нейронів, а також відео процесу повної синхронізації невеликої мережі (відповідно до передбачення MSF) наведено праворуч. олові" аплізіі реагує сильним втягуванням зябра (що можна розглядати як аналог безумовного рефлексу). Після низки спільних впливів тварина реагує на слабке дотик ксіфону таким же активним втягуванням зябра, як і у відповідь на удар струмом по "голові". Упрошенная схема ділянки нейронної мережі, що реалізує таку поведінку, наведена на рис. 4.4. На рис. 4.4, А представлений варіант, при якому асоціативне навчання реалізується на рівні контактів нейронів трьох типів: нейрона - датчика торкань сифона (/), нейрона - датчика больового впливу (3) і нейрона, що запускає руховий акт стиснення м'язи мантії (2). На рис. 4.4, Б розглянуто більш складний варіант, при якому в схему включений додатковий елемент - інтернейрон (4), що грає роль прообразу ділянки нейронної мережі, що володіє неко- Р і с. 4.4. Схема двох варіантів утворення умовного рефлексу (асоціативного зв'язку) в ділянці нейронної мережі морського равлика аплізіі (Aplisia punctata). В обох варіантах ланцюг безумовного рефлексу: роздратування датчика болю (нейрон 3) - стиснення м'язи (нейрон 2). А - спрощений варіант: 6 - варіант з використанням вставних нейронів (інтернейронов). На вставці показана схема синаптического з'єднання, де (а) прс-.а (б) постеінаптіческіс закінчення нейронів, гуртки - молекули нейромедіатора. стрілка вказує напрям проходження сигналу торою інтелектуальною функцією. Групи інтернейронов або вставних нейронів, тобто нейронів, вставлених між сенсорними і руховими нейронами, дійсно стають все більш і більш сильно розвиненими в міру просування тварин по філогенетичної сходах. Схема першого упрошенной варіанти формування ланцюга умовного рефлексу така. Спочатку торкання сифона не веде до реакції втягування зябра, але після кількох десятків поєднань торкання сифона і подальшого удару струмом по голові з'являється умовний рефлекс. Механізм рефлексу полягає в тому, що активація нейрона 3 полегшує проведення сигналу через синапси між нейронами / і 2. У другому варіанті механізм рефлексу полягає в активації інтернейрон 4, закінчення якого збільшують локальний електричний потенціал в певному районі "командного" нейрона 2. У підсумку цього досить складного процесу відбувається зміна геометрії в області синаптичних з'єднань відростків інтернейрон 4, командного нейрона 2 і нейрона-датчика торкань /. ^ Го зміна пов'язана із збільшенням розмірів так званих "шипиків", що і являє собою матеріальну реалізацію процесу навчання. Місця з'єднання нейронів - синапси, або синаптичні щілини, - відіграють важливу роль у механізмах навчання. Структуру цих щілин утворюють пре-і постсинаптичні нейронні мембрани, розділені міжнейронних простором. Нервовий імпульс (або, як його часто називають, потенціал дії) передається від пресинаптичної мембрани одного нейрона до постсинаптичні мембрані іншого. При цьому сам процес передачі являє собою складне, багатоетапне подія, що реалізовується за допомогою роботи електричних і молекулярних механізмів. У результаті проходження нервових імпульсів по "вхідному", пресинаптичними нейрона в районі синаптичної щілини із спеціалізованих структур пресинаптичних закінчень виділяються молекули нейромедіатора, які перетинають синаптичну щілину, активують рецептори постсинаптичної мембрани і викликають появу по-стсінаптіческіх потенціалів, зростання "шипиків", зміна геометрії ділянок мережі і в кінцевому рахунку запам'ятовування і асоціативне навчання. На підставі експериментальних даних останніх десятиліть стає ясно, що матеріальним субстратом закріплення асоціативних зв'язків, зокрема і запам'ятовування взагалі. є саме синаптичні структури. У результаті утворення асоціації, запам'ятовування або фіксації будь-якої інформації відбувається формування нової конфігурації нейронних зв'язків ділянок нейронних мереж. Мережеві структури формують сязасчеттого, що отелення кожного нейрона (нервової клітини) відходить безліч гілочок дендритів, які отримують вхідні сигнали від інших клітин, і один тонкий розгалужених аксон, по якому сигнал після обробки передається до інших нервовим клітинам. При такій будові за рахунок свого ветвящегося дендритного дерева кожен нейрон може одержувати сигнали від безлічі інших нервових клітин. Оброблений сигнал також може бути переданий багатьом нейронам за рахунок розгалужених кінцевих закінчень аксона. Матеріали взяті з: http://nebook.net/book_psihologya-pedagogka_626_page_45
ВЧЕННЯ І.П. ПАВЛОВА ПРО ВИЩУ НЕРВОВУ ДІЯЛЬНІСТЬ.
Поиск по сайту: |












 ) кожного з N нейронів задається наступним чином:
) кожного з N нейронів задається наступним чином: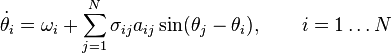
 це частота коливання i-того осцилятору,
це частота коливання i-того осцилятору,  та
та  — сила з'єднання та елемент матриці з'єднань відповідно.
— сила з'єднання та елемент матриці з'єднань відповідно. , то кажуть що мережа з N нейронів повністю синхронізована, якщо її стан відповідає рівнянню
, то кажуть що мережа з N нейронів повністю синхронізована, якщо її стан відповідає рівнянню  . Стан повної синхронізації може бути досягнутий тілки у мережі з ідентичними нейронами.
. Стан повної синхронізації може бути досягнутий тілки у мережі з ідентичними нейронами.


 ;
;  це D-вимірний вектор змінних i-того нейрона (вектор стану);
це D-вимірний вектор змінних i-того нейрона (вектор стану);  та
та  — векторні поля що описують відповідно (нелінійну) внутрішню динаміку (звичайне мат. визначення нейрону) та функцію зв'язку між елементами;
— векторні поля що описують відповідно (нелінійну) внутрішню динаміку (звичайне мат. визначення нейрону) та функцію зв'язку між елементами;  елемент матриці
елемент матриці  що містить сили синаптичних зв'язків між нейронами.
що містить сили синаптичних зв'язків між нейронами. . Часто потрібно аналізувати мережу з великою кількістю нейронів (
. Часто потрібно аналізувати мережу з великою кількістю нейронів (  ), і тоді прямий аналіз стабільності синхронізованого стану мережі (за допомогою, наприклад, експонент Ляпунова) стає занадто громіздкою обчислювальною задачею. З іншого боку, відповідно до формалізму MSF, для визначення стабільності синхронізованого стану вищенаведеної системи, достатньо розрахувати експоненти Ляпунова для набагато меншої, лінеаризованої D-вимірної системи, що задається рівнянням (припустимо, що матриця зв'язків має виключно реальні власні числа):
), і тоді прямий аналіз стабільності синхронізованого стану мережі (за допомогою, наприклад, експонент Ляпунова) стає занадто громіздкою обчислювальною задачею. З іншого боку, відповідно до формалізму MSF, для визначення стабільності синхронізованого стану вищенаведеної системи, достатньо розрахувати експоненти Ляпунова для набагато меншої, лінеаризованої D-вимірної системи, що задається рівнянням (припустимо, що матриця зв'язків має виключно реальні власні числа):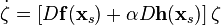
 — якобіан векторного поля, розрахований уздовж синхронізованого рішення
— якобіан векторного поля, розрахований уздовж синхронізованого рішення 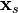 (під синхронізованим рішенням тут мається на увазі чисельне рішення рівняння одного нейрона, з'єднаного з самим собою), а
(під синхронізованим рішенням тут мається на увазі чисельне рішення рівняння одного нейрона, з'єднаного з самим собою), а  — спеціальний скалярний параметр.
— спеціальний скалярний параметр. ) невеликої лінійної системи (2), для потрібного діапазону параметру
) невеликої лінійної системи (2), для потрібного діапазону параметру 