|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Подробное описание алгоритма
Отбор (селекция) На этапе отбора нужно из всей популяции выбрать определённую её долю, которая останется «в живых» на этом этапе эволюции. Есть разные способы проводить отбор. Вероятность выживания особи h должна зависеть от значения функции приспособленности Fitness(h). Сама доля выживших s обычно является параметром генетического алгоритма, и её просто задают заранее. По итогам отбора из N особей популяции H должны остаться sN особей, которые войдут в итоговую популяцию H'. Остальные особи погибают. · Турнирная селекция - сначала случайно выбирается установленное количество особей (обычно две), а затем из них выбирается особь с лучшим значением функции приспособленности · Метод рулетки - вероятность выбора особи тем вероятнее, чем лучше её значение функции приспособленности · Метод ранжирования - вероятность выбора зависит от места в списке особей отсортированном по значению функции приспособленности · Равномерное ранжирование - вероятность выбора особи определяется выражением: · Сигма отсечение - для предотвращения преждевременной сходимости генетического алгоритма используются методы масштабирующие значение целевой функции. Вероятность выбора особи тем больше, чем оптимальнее значение масштабируемой целевой функции Выбор родителей Размножение в генетических алгоритмах обычно половое — чтобы произвести потомка, нужны несколько родителей, обычно два. Можно выделить несколько операторов выбора родителей: 1. Панмиксия - оба родителя выбираются случайно, каждая особь популяции имеет равные шансы быть выбранной 2. Инбридинг - первый родитель выбирается случайно, а вторым выбирается такой, который наиболее похож на первого родителя 3. Аутбридинг - первый родитель выбирается случайно, а вторым выбирается такой, который наиболее не похож на первого родителя Инбридинг и аутбридинг бывают в двух формах: фенотипной и генотипной. В случае фенотипной формы похожесть измеряется в зависимости от значения функции приспособленности (чем ближе значения целевой функции, тем особи похожее), а в случае генотипной формы похожесть измеряется в зависимости от представления генотипа (чем меньше отличий между генотипами особей, тем особи похожее). Размножение (Скрещивание) Размножение в разных алгоритмах определяется по-разному — оно, конечно, зависит от представления данных. Главное требование к размножению — чтобы потомок или потомки имели возможность унаследовать черты обоих родителей, «смешав» их каким-либо способом. Почему особи для размножения обычно выбираются из всей популяции H, а не из выживших на первом шаге элементов H0 (хотя последний вариант тоже имеет право на существование)? Дело в том, что главный бич многих генетических алгоритмов — недостаток разнообразия (diversity) в особях. Достаточно быстро выделяется один-единственный генотип, который представляет собой локальный максимум, а затем все элементы популяции проигрывают ему отбор, и вся популяция «забивается» копиями этой особи. Есть разные способы борьбы с таким нежелательным эффектом; один из них — выбор для размножения не самых приспособленных, но вообще всех особей. Мутации К мутациям относится все то же самое, что и к размножению: есть некоторая доля мутантов m, являющаяся параметром генетического алгоритма, и на шаге мутаций нужно выбрать mN особей, а затем изменить их в соответствии с заранее определёнными операциями мутации. Критика Существует несколько поводов для критики насчёт использования генетического алгоритма по сравнению с другими методами оптимизации: · Повторная оценка функции приспособленности (фитнесс-функции) для сложных проблем, часто является фактором, ограничивающим использование алгоритмов искусственной эволюции. Поиск оптимального решения для сложной задачи высокой размерности зачастую требует очень затратной оценки функции приспособленности. В реальных задачах, таких как задачи структурной оптимизации, единственный запуск функциональной оценки требует от нескольких часов до нескольких дней для произведения необходимых вычислений. Стандартные методы оптимизации не могут справиться с проблемами такого рода. В таком случае, может быть необходимо пренебречь точной оценкой и использовать аппроксимацию пригодности, которая способна быть вычислена эффективно. Очевидно, что применение аппроксимации пригодности может стать одним из наиболее многообещающих подходов, позволяющих обоснованно решать сложные задачи реальной жизни с помощью генетических алгоритмов. · Генетические алгоритмы плохо масштабируемы под сложность решаемой проблемы. Это значит, что число элементов, подверженных мутации очень велико, если велик размер области поиска решений. Это делает использование данной вычислительной техники чрезвычайно сложным при решении таких проблем, как, например, проектирование двигателя, дома или самолёта. Для того чтобы сделать так, чтобы такие проблемы поддавались эволюционным алгоритмам, они должны быть разделены на простейшие представления данных проблем. Таким образом, эволюционные вычисления используются, например, при разработке формы лопастей, вместо всего двигателя, формы здания, вместо подробного строительного проекта и формы фюзеляжа, вместо разработки вида всего самолёта. Вторая проблема, связанная со сложностью, кроется в том, как защитить части, которые эволюционировали с высокопригодными решениями от дальнейшей разрушительной мутации, в частности тогда, когда от них требуется хорошая совместимость с другими частями в процессе оценки пригодности. Некоторыми разработчиками было предложено, что подход, предполагающий развитие пригодности эволюционирующих решений, смог бы преодолеть ряд проблем с защитой, но данный вопрос всё ещё остаётся открытым для исследования. · Решение является более пригодным лишь по сравнению с другими решениями. В результате условие остановки алгоритма неясно для каждой проблемы. · Во многих задачах генетические алгоритмы имеют тенденцию сходиться к локальному оптимуму или даже к спорным точкам, вместо глобального оптимума для данной задачи. Это значит, что они «не знают», каким образом пожертвовать кратковременной высокой пригодностью для достижения долгосрочной пригодности. Вероятность этого зависит от формы ландшафта пригодности: отдельные проблемы могут иметь выраженное направление к глобальному минимуму, в то время как остальные могут указывать направление для фитнесс-функции на локальный оптимум. Эту проблему можно решить использованием иной фитнесс-функции, увеличением вероятности мутаций, или использованием методов отбора, которые поддерживают разнообразие решений в популяции, хотя Теорема об отсутствии бесплатного обеда при поиске и оптимизации[15] доказывает, что не существует общего решения данной проблемы. Общепринятым методом поддержания популяционного разнообразия является установка уровневого ограничения на численность элементов с высоким сродством, которое снизит число представителей сходных решений в последующих поколениях, позволяя другим, менее сходным элементам оставаться в популяции. Данный приём, тем не менее, может не увенчаться успехом в зависимости от ландшафта конкретной проблемы. Другим возможным методом может служить простое замещение части популяции случайно сгенерированными элементами, в момент, когда элементы популяции становятся слишком сходны между собой. Разнообразие важно для генетических алгоритмов (и генетического программирования) потому, что перекрёст генов в гомогенной популяции не несёт новых решений. В эволюционных стратегиях и эволюционном программировании, разнообразие не является необходимостью, так как большая роль в них отведена мутации. Классический генетический алгоритм (также называемый элементарным или простым генетическим алгоритмом) состоит из следующих шагов:
Рассмотрим первые три этапа: инициализация, оценка приспособленности хромосом и проверка условия остановки алгоритма. Инициализация, т.е. формирование исходной популяции. Каждая особь популяции представляет собой решение задачи, иначе говоря, является кандидатом на решение. Формирование исходной популяции, как правило, происходит с использованием какого-нибудь случайного закона, в ряде случаев исходная популяция может быть также результатом работы другого алгоритма. Необходимо отметить, что особи популяции могут состоять как из одной, так и из нескольких хромосом. Каждая хромосома особи в свою очередь состоит из генов, причем количество генов в хромосоме определяется количеством варьируемых параметров решаемой задачи (аргументов целевой функции). Для применения генетических операторов значения этих параметров должны быть представлены в виде двоичной последовательности, т.е. двоичной строки, состоящей из нескольких бит. Количество бит при кодировании гена (хромосомы) зависит от требуемой точности решаемой задачи. Оценивание приспособленности хромосом в популяции состоит в расчете функции приспособленности для каждой хромосомы этой популяции. Чем больше значение этой функции, тем выше «качество» хромосомы. Форма функции приспособленности зависит от характера решаемой задачи. Предполагается, что функция приспособленности всегда принимает неотрицательные значения и, кроме того, что для решения оптимизационной задачи требуется максимизировать эту функцию. Если исходная форма функции приспособленности не удовлетворяет этим условиям, то выполняется соответствующее преобразование (например, задачу минимизации функции можно легко свести к задаче максимизации). Проверка условия остановки алгоритма. Определение условия остановки генетического алгоритма зависит от его конкретного применения. В оптимизационных задачах, если известно максимальное (или минимальное) значение функции приспособленности, то остановка алгоритма может произойти после достижения ожидаемого оптимального значения, возможно – с заданной точностью. Остановка алгоритма также может произойти в случае, когда его выполнение не приводит к улучшению уже достигнутого значения. Генетический алгоритм может быть также остановлен по истечении определенного времени выполнения либо после выполнения заданного количества итераций. Если условие остановки выполнено, то производится переход к завершающему этапу выбора «наилучшей» хромосомы. В противном случае на следующем шаге выполняется селекция. Применение генетических алгоритмов Генетические алгоритмы применяются для решения следующих задач: 1. Оптимизация функций 2. Оптимизация запросов в базах данных 3. Разнообразные задачи на графах (задача коммивояжёра, раскраска, нахождение паросочетаний) 4. Настройка и обучение искусственной нейронной сети 5. Задачи компоновки 6. Составление расписаний 7. Игровые стратегии 8. Теория приближений 9. Искусственная жизнь 10. Биоинформатика (свёртывание белков) Подробное описание алгоритма Оператор скрещивания. На первом этапе скрещивания выбираются пары хромосом из родительской популяции (родительского пула). Это временная популяция, состоящая из хромосом, отобранных в результате селекции и предназначенных для дальнейших преобразований операторами скрещивания и мутации с целью формирования новой популяции потомков. На данном этапе хромосомы из родительской популяции объединяются в пары. Это производится случайным способом в соответствии с вероятностью скрещивания 1. потомок, хромосома которого на позициях от 2. потомок, хромосома которого на позициях от Действие оператора скрещивания проиллюстрировано следующим примером.
Оператор мутации с вероятностью
Как уже упоминалось выше, вероятность мутации обычно очень мала, и именно от нее зависит, будет данный ген мутировать или нет. Вероятность Формирование новой популяции. Хромосомы, полученные в результате применения генетических операторов к хромосомам временной родительской популяции, включаются в состав новой популяции. Она становится так называемой текущей популяцией для данной итерации генетического алгоритма. На каждой очередной итерации рассчитываются значения функции приспособленности для всех хромосом этой популяции, после чего проверяется условие остановки алгоритма и либо фиксируется результат в виде хромосомы с наибольшим значением функции приспособленности, либо осуществляется переход к следующему шагу генетического алгоритма, т.е. к селекции. В классическом генетическом алгоритме вся предшествующая популяция хромосом замещается новой популяцией потомков, имеющей ту же численность.
Поиск по сайту: |
 , где
, где 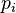 - вероятность выбора i особи,
- вероятность выбора i особи,  - значение функции приспособленности для i особи,
- значение функции приспособленности для i особи, 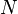 - количество особей в популяции
- количество особей в популяции , где
, где  ,
,  ,
,  - порядковый номер особи в списке особей отсортированном по значению функции приспособленности ( т.е.
- порядковый номер особи в списке особей отсортированном по значению функции приспособленности ( т.е.  - если мы минимизируем значение функции приспособленности)
- если мы минимизируем значение функции приспособленности) , где
, где 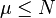 параметр метода
параметр метода , где
, где  ,
,  - среднее значение целевой функции для всей популяции, σ - среднеквадратичное отклонение значения целевой функции
- среднее значение целевой функции для всей популяции, σ - среднеквадратичное отклонение значения целевой функции . Далее для каждой пары отобранных таким образом родителей разыгрывается позиция гена (локус) в хромосоме, определяющая так называемую точку скрещивания. Если хромосома каждого из родителей состоит из
. Далее для каждой пары отобранных таким образом родителей разыгрывается позиция гена (локус) в хромосоме, определяющая так называемую точку скрещивания. Если хромосома каждого из родителей состоит из  генов, то очевидно, что точка скрещивания
генов, то очевидно, что точка скрещивания  представляет собой натуральное число, меньшее
представляет собой натуральное число, меньшее  . В результате скрещивания пары родительских хромосом получается следующая пара потомков:
. В результате скрещивания пары родительских хромосом получается следующая пара потомков: до
до  до
до 
 , изменяет значение гена в хромосоме на противоположное (т.е. с
, изменяет значение гена в хромосоме на противоположное (т.е. с  на
на 
 для каждого гена и отбором для выполнения этой операции тех генов, для которых разыгранное число оказывается меньшим или равным значению
для каждого гена и отбором для выполнения этой операции тех генов, для которых разыгранное число оказывается меньшим или равным значению