|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Паратгормон. Роль в обмене веществСтр 1 из 2Следующая ⇒
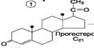
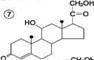
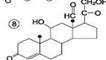
Механизм клеточного действия гормонов. С участием ц-АМФ. Механизм с прямым взаимодействием с хроматином. Гор. прямого действия. К этой группе относятся липофильные гор. – стероиды и йодтиронины. они мало растворимы в воде и поэтому образуют в крови комплексные соединения с белками плазмы. Гормоны прямого действия в силу своей липофильности способны диффундировать через двойной липидный слой мембран клеток-миш.. Рецепторы к этим гормонам находятся в цитозоле. Образующийся комплекс гормона с рецеп. перемещается в ядро клетки, где связывается с хроматином и воздействует на ДНК. В результате изменяется скорость синтеза РНК на матрице ДНК (транскрипция) и скорость образования специфических ферментативных белков на матрице РНК (трансляция). Это приводит к изменению кол-ва ферментативных белков в клетках-миш. и изменению в них направленности хим. реакций . Гормоны дистантного действия. К ним относятся гидрофильные гор – катехоламины и гормоны белково-пептидной природы. Так как эти в-ва не растворимы в липидах, они не могут проникать через клеточные мембраны. Рецепторы для этих гормонов расположены на наруж. Пов. Плазм. мембраны клеток-миш. Гор. реализуют своё действие на клетку при помощи вторичного посредника, в качестве которого чаще всего цАМФ. Взаимодействие гор. с его специфическим рецептором приводит к активации G-белка клеточной мембраны. G-белок связывает ГТФ и активирует аденилатциклазу. Активная аденилатциклаза превращает АТФ в цАМФ, цАМФ активирует протеинкиназу. Неак. протеинкиназа представляет собой тетрамер, который состоит из 2 регуляторных (R) и 2 каталитических (C) субъед. В результате взаимодействия с цАМФ происходит диссоц. тетрамера и освобождается активный центр фер. Протеинкиназа фосфорилирует белки-ферменты за счёт АТФ, либо активируя их, либо инактивируя. В результате этого изменяется (увеличивается или уменьшается) скорость химических реакций в клетках-миш. Гор. ЩИТ. Ж. Роль тироксинаи тиреокальцитонина в обмене веществ. йодтиронины (трийодтиронин, Т3, тетрайодтиронин Т4) Биосинтез Йодтиронины синтезируются в составе белка тиреоглобулина в фолликулах щит. ж. Тиреоглобулин синтезируется на риб. шер. ЭР в виде претиреоглобулина, затем переносится в цистерны ЭР откуда поступает в аппарат Гольджи, включается в состав секреторных гранул и секретируется во внеклеточный коллоид, где происходит йодирование остатков тирозина и образование йодтиронинов. Что и осуществляется в несколько этапов: Транспорт йода в клетки щитовидной железы.Йод поступает в ЖКТ с пищей и водой. Транспорт йодида в клетки щитовидной железы - энергозависимый процесс и происходит при участии специального транспортного белка Работа этого йодид-переносящего белка сопряжена с Nа+,К+-АТФ-азой.Окисление йода. происходит при участии тиреопероксидазы и Н2О2 в качестве окислителяЙодирование тирозина.Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина. катализатор тиреопероксидазой. Образование йодтиронинов. окисленный йод реагирует с остатками тирозина с образованием монойод-тирозинов (МИТ) и дийодтирозинов (ДИТ). Две молекулы ДИТ конденсируются с образованием йодтиронина Т4, а МИТ и ДИТ - с образованием йодтиронина Т3. Йодтиреоглобулин транспортируется из коллоида в фолликулярную клетку путём эндоцитоза и гидролизуется ферментами лизосом с освобождением Т3 и Т4. РОЛЬ 1 ускорении белкового синтеза, стимуляции процессов роста и клеточной дифференцировки.2Т3 ускоряет транскрипцию гена гормона роста. 4 Т3 стимулирует работу Na+,К+-АТФ-азы 5 ускоряют гликолиз, липолиз синтез холестерола и жёлчных кислоти. тиреокальцитонин является антагонистом паратгормона вызывающий понижение содержания Ca2+ в плазме крови Паратгормон. Роль в обмене веществ. Паратгормон - одноцепочечный полипептид действие которого направлено на повышение концентрации ионов кальция и снижение концентрации фосфатов в плазме крови.Органы-мишенидля ПТГ - кости и почки.1 стимулирует мобилизацию кальция из кости; 2 - стимулирует реабсорбцию ионов кальция в дистальных канальцах почек; 3 - активирует образование кальцитриола, 4 повышает концентрацию кальция в межклеточной жидкости, 5 В почках стимулирует реабсорбцию кальция в дистальных извитых канальцах и тем самым снижает экскрецию кальция с мочой, 6 уменьшает реабсорбцию фосфатов. индуцирует синтез кальцитриола Гормоны коры надпочеч. Минералкортикоиды биосинтез и строение, роль в обмене веществ. В коре надпочеч.синтезируется Глюкокортикоиды, Минералокортикоиды, Андрогены Синтез
Активация канальцевой реабсорбции натрия из мочи в эпителиальные клетки канальцев и далее в межклеточное пространство, а калий наоборот экскретируется, вместе с ионами водорода, с мочой; Поддержание осмотического давления, АД, возбудимости клетки; Регуляция ионного транспорта в потовых, слюнных железах и ЖКТ; При избытке: гиперволемия, гипертензия, алкалоз, нарушение сердечного ритма, При недостатке: гиповолемия, гиперкалиемия, гипотензия, нарушение сердечного ритма, мозговые нарушения, избыток вазопрессина (АДГ), за счет экскреции воды с ионами натрия, нарушение пищеварения
Нарушение обмена веществ при инсулиновой недостаточности.При недостаточном синтезе инсулина развивается специфическое заболевание –сахарный диабет Помимо клинически выявляемых симптомов (полиурия, полидипсия и полифагия), сахарный диабет характеризуется рядом специфических нарушений процессов обмена. Так, у больных развиваются гипергликемия (увеличение уровня глюкозы в крови) и гликозурия (выделение глюкозы с мочой, в которой в норме она отсутствует). К расстройствам обмена относят также усиленный распад гликогена в печени и мышцах, замедление биосинтеза белков и жиров, снижение скорости окисления глюкозы в тканях, развитие отрицательного азотистого баланса, увеличение содержания холестерина и других липидов в крови. При диабете усиливаются мобилизация жиров из депо, синтез углеводов из аминокислот (глюконеогенез) и избыточный синтез кетоновых тел (кетонурия). После введения больным инсулина все перечисленные нарушения, как правило, исчезают, однако действие гормона ограничено во времени, поэтому необходимо вводить его постоянно. Клинические симптомы и метаболические нарушения при сахарном диабете могут быть объяснены не только отсутствием синтеза инсулина Клеточный механизм действия инсулина на уровне органов мишеней. Действие инсулина начинается с его связывания со специфическим рецеп. на поверхности клетки-мишени. Инсулиновый рецептор представляет собой сложный интегральный белок клеточной мембраны, построенный из 2 субъединиц (a и b). Инсулин с высокой специфичностью связывается и распознаётся а-субъединицей рецептора, которая при присоединении гормона изменяет свою конформацию. Это приводит к появлению тирозинкиназной активности у субъединицы b, что запускает разветвлённую цепь реакций по активации ферментов, которая начинается с аутофосфорилирования рецептора. Это приводит к фосфорилиро-ванию других внутриклеточных белков - субстратов инсулинового рецептора (IRS). Известно несколько таких субстратов: IRS-1, IRS-2 все Это приводит к активации нескольких сигнальных путей, представляющих каскад реакций активации специфических протеинкиназ. В результате активации протеинкиназ происходит фосфорилирование ферментов и факторов транскрипции, что составляет основу многочисленных эффектов инсулина. Белок, известный как Ras-белок в неактивном состоянии прикреплён к внутренней поверхности плазматической мембраны и связан с ГДФ. Стимуляция инсулином приводит к образованию активной ГТФ-связан-ной формы Ras. Один из субстратов инсулинового рецептора Shc участвует в образовании комплекса с небольшим цитозольным белком Grb. Образовавшийся комплекс взаимодействует с Ras-белком. В этот комплекс включаются другие белки. Они способствуют отделению ГДФ от Ras-белка и присоединению ГТФ. Активированный Ras соединяется с протеинкиназой Raf-1. При участии Raf-1 сначала фосфорилируется и активируется киназа МАПК, которая, в свою очередь, фос-форилирует МАПК.
Глюкагон, рольв обмене веществ. Эффекты глюкагонав основном противоположны эффектам инсулина. Основные клетки-мишени глюкагона - печень и жировая ткань. Связываясь с рецепторами на плазматической мембране клеток-мишеней, глюкагон повышает содержание цАМФ В гепатоци-тах это приводит к активации фосфорилазы гликогена и к снижению активности гликоген-синтазы. В результате ускоряется мобилизация гликогена. Кроме того, глюкагон стимулирует глюконеогенез, индуцируя синтез ферментов: глюкозо-6-фосфатазы, фосфоенол-пируваткарбоксикиназы, фруктозо-1,6-бис-фосфатазы В клетках жировой ткани глюкагон через аденилатциклазный каскад активирует гормончувствительную ТАГ-липазу и стимулирует липолиз Таким образом, в противоположность инсулину глюкагон стимулирует мобилизацию основных энергоносителей - углеводов и жиров.
Поиск по сайту: |



 Основные эффекты минералокортикоидов:
Основные эффекты минералокортикоидов: