|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Флористическое районирование земного шара
Флористическим районированием называют разделение поверхности земного шара на флористические регионы различного ранга (фитохорионы, или фитохории). Каждый регион представляет собой территорию, в пределах которой флора более или менее однородна, однотипна, имеет свою специфику и в большей или меньшей степени отличается от флоры других регионов. При флористическом районировании выделяют территориальные единицы различного ранга, которые находятся в строгом соподчинении. Самой крупной единицей флористического районирования является царство. На земном шаре выделяют шесть царств: Голарктическое (Holarctis), Палеотропическое (Palaeotropis), Неотропическое (Neotropis), Капское (Capensis), Австралийское (Australis) и Голантарктическое (Antarctis) (рис. 15.18 ). Флористические царства подразделяют на области, области - на провинции, провинции - на округа. Флористические округа можно подразделять на флористические районы, а последние - на районы конкретных флор. Для царств характерны эндемичные семейства, и очень высок родовой и видовой эндемизм. Области, как правило, эндемичных семейств не имеют, но степень родового и видового эндемизма очень высока. Во флорах провинций и округов преобладают эндемичные виды. Различия во флоре соседних флористических районов еще меньше. Необходимо отметить, что четких границ между отдельными фитохорионами в природе обычно нет, и проводить их приходится более или менее условно. Состав флор меняется в пространстве то быстрее, то медленнее, но чаще всего постепенно, нерезко. Флористические царства представляют собой территории, очень сильно различающиеся одна от другой по флоре. Эти различия нельзя объяснить какими-то современными факторами, например климатическими условиями. Решающую роль в формировании флоры того или иного царства играли исторические факторы.
Рис. 15.18. Флористические царства Земли : I – Голарктическое; II – Палеотропическое; III – Неотропическое; IV – Капское; V – Австралийское; VI - Голанктарктическое. Голарктическое царство - самое обширное по площади. Оно занимает больше половины всей суши земного шара и охватывает всю внетропическую часть Северного полушария. Однако голарктическая флора довольно бедна - всего около 30 тыс. видов. В Голарктическом царстве насчитывается более 30 эндемичных семейств растений (гинкговые, эвкоммиевые, пионовые, диапенсиевые, адоксовые, сусаковые и др.). Однако все эти семейства мелкие, весьма малочисленные по числу представителей, нередко включающие только один вид. Гораздо лучше характеризуют флору Голарктического царства некоторые, хотя и не эндемичные, но широко распространенные здесь семейства. Подавляющее большинство видов этих семейств приурочено именно к Голарктису. К таким семействам относятся лютиковые, березовые, гвоздичные, маревые, гречишные, ивовые, крестоцветные, зонтичные и ряд других. Из голосеменных следует назвать семейства сосновых и кипарисовых. Во флоре Голарктического царства насчитывается очень много эндемичных родов и видов. Голарктическое царство подразделяется на три подцарства. Самым большим по площади и богатым во флористическом отношении является Бореальное подцарство. В его состав входит почти вся Европа, значительная часть Азии и Северной Америки. В пределах подцарства выделяют четыре области. Территория России целиком входит в наиболее крупную из них Циркумбореальную область. В ее флоре нет эндемичных семейств, а эндемичных родов сравнительно немного. Характерными родами, имеющими широкое распространение в данной области, являются дуб (Quercus), береза (Betula), ольха (Alnus), клен (Acer), тополь (Populus), ива (Salix), боярышник (Crataegus), груша (Pyrus), яблоня (Malus), рябина (Sorbus) и многие другие. Палеотропическое царство занимает огромную территорию в тропической области Старого Света и уступает по площади только Голарктическому. Оно включает Африку (кроме северной части и крайнего юга), остров Мадагаскар, Индию, Индокитай, многочисленные крупные и мелкие острова между Азией и Австралией, многие островные территории в Тихом океане. Таким образом, территория царства сильно расчленена, а отдельные его части далеко разбросаны в пределах тропического пояса - от Африки до Полинезии. Тем не менее, флора разных районов царства имеет много общего. Прежде всего, необходимо отметить исключительное богатство флоры рассматриваемой территории. Здесь встречаются многие десятки тысяч видов. Флора царства достаточно своеобразна, насчитывает около 40 эндемичных семейств (непентесовые, банановые, панданусовые и др.). Число эндемичных родов и особенно видов настолько велико, что не поддается точному учету. В рассматриваемом царстве богато представлено семейство тутовых, особенно род фикус (Ficus), насчитывающий более 1000 видов. Широко распространены семейства диптерокарповых, бальзаминовых, молочайных, аралиевых, ароидных, имбирных и др. Характерны для Палеотрописа некоторые роды пальм, например борассус (Borassus), гифене (Hyphaene), рафия (Raphia) и др. Встречаются многие роды саговниковых - очень древних голосеменных растений, по внешнему облику несколько напоминающих пальмы. Палеотропическое царство подразделяют на пять подцарств. Неотропическое царство включает преобладающую часть Южной Америки (к северу от 30° ю. ш.), Центральную Америку вплоть до юга Мексики, острова Карибского моря и южную оконечность полуострова Флорида. Неотропическое царство довольно обширно по площади, но все же уступает в этом отношении Палеотропису и тем более Голарктису. Флора царства необычайно богата и по числу видов сопоставима только с флорой Палеотрописа. Здесь насчитывается много десятков тысяч видов, принадлежащих к нескольким сотням семейств. Точное число видов неизвестно. О видовом богатстве царства можно судить хотя бы по тому, что флора только одной Бразилии содержит более 40 000 видов. Флора Неотрописа отличается большим своеобразием. Здесь насчитывается около 25 эндемичных семейств, огромное количество эндемичных родов и особенно видов. В Неотропическом царстве широко распространены такие семейства, как настурциевые, кактусовые, синюховые, ароидные, бромелиевые и др. Именно в Неотрописе встречается в естественных сообществах подавляющее большинство представителей семейства кактусовых. К числу характерных для Неотрописа относится род бегония (Begonia), представленный здесь очень многими видами, а также роды фуксия (Fuchsia), юкка (Yucca), агава (Agave). В Неотрописе имеются и свои особые роды пальм, например хамедорея (Chamaedorea), маурития (Mauritia), сабаль (Sabal) и др. Капское царство - самое маленькое по площади. Оно занимает небольшую территорию на крайнем юге Африки. Однако флора его исключительно богата (около 7000 видов) и своеобразна. Она резко отличается от флоры соседних территорий, расположенных к северу. В Капском флористическом царстве насчитывается семь эндемичных семейств, эндемичных родов более 210. Эндемичен для Капского царства и род амариллис (Amaryllis), куда относятся знакомые многим комнатные растения. Интересно отметить, что имеется 14 эндемичных родов из очень древнего семейства протейных (основная масса видов этого семейства сосредоточена в Австралии). Особенно богаты видами такие эндемичные роды, как протея (Protea) и леукадендрон (Leucadendron). Большое видовое разнообразие наблюдается также в некоторых родах, которые не являются эндемичными и относятся к разным семействам. Так, в роде вереск (Erica) насчитывается свыше 600 видов, в роде пеларгониум (Pelargonium) - более 200. Характерно обилие видов, относящихся к семействам амариллисовых, ирисовых и рестионовых. Своеобразие, самобытность флоры Капского царства объясняется тем, что растительный мир данной территории длительное время развивался в условиях изоляции. Преградой для обмена флорой с остальной частью Африканского континента в настоящее время служат пустыни, расположенные к северу от Капского царства. Именно они препятствуют смешению капской флоры с соседними флорами. Австралийское царство включает Австралию и соседний остров Тасмания, а также некоторые мелкие острова. Австралийское царство занимает совершенно изолированное положение. Оно отделено от остальной суши более или менее обширными морскими просторами. Территория царства довольно велика, по размерам она лишь немногим меньше Европы. Флора Австралийского царства очень богата (около 15 000 видов), чрезвычайно самобытна, оригинальна и содержит множество древних растений. Австралийская флора отличается очень высоким эндемизмом на всех уровнях. Здесь имеется более десятка эндемичных семейств, эндемичных родов насчитывается около 570. Доля эндемичных видов в целом достигает 75-80%, в некоторых районах эта цифра еще выше. В составе австралийской флоры немало характерных семейств. Среди них нужно назвать в первую очередь семейство протейных. Большинство видов этого семейства (свыше 700) сосредоточены именно здесь. Наиболее богатые видами роды протейных - гревиллея (Grevillea), хакея (Hakea), банксия (Banksia). К числу характерных для австралийской флоры относится семейство казуариновых. Оно включает деревья своеобразного облика, несколько напоминающие по внешнему виду хвойные. Но все же самыми характерными для австралийской флоры следует считать эвкалипты и акации. Эвкалиптов насчитывается здесь более 600 видов (род Eucalyptus относится к семейству миртовых). Большинство их - деревья, но есть также и кустарники. Почти все эвкалипты - вечнозеленые растения. Акации (Acacia), как и эвкалипты, весьма многочисленны (более 500 видов) и очень разнообразны. Это вечнозеленые деревья и кустарники. У многих акаций вместо настоящих листьев развиваются филлодии - плоские зеленые черешки разнообразной формы. Одна из примечательных особенностей флоры Австралийского царства - отсутствие некоторых широко распространенных на других континентах семейств растений и даже более крупных таксонов. Здесь нет, например, хвощей, бамбуков, представителей подсемейства яблоневых семейства розоцветных. Отсутствуют семейства вересковых, бегониевых, валериановых, чайных. Это явление иногда обозначают термином “дефектность флоры”. Голантарктическое царство расположено в Южном полушарии, причем в относительно высоких широтах. Оно включает южную часть Южной Америки (примерно от 30° ю. ш.), Новую Зеландию, незначительные, не покрытые льдами, участки Антарктиды и многочисленные мелкие острова, находящиеся в южной части Атлантического, Индийского и Тихого океанов. Территория царства сравнительно невелика и чрезвычайно сильно раздроблена. Отдельные части царства расположены очень далеко друг от друга, разделены огромными морскими просторами. Тем не менее, флора всех этих участков суши имеет явные черты сходства. Есть много общих родов и даже видов. В качестве примера таких родов можно назвать колобантус (Colobanthus) из семейства гвоздичных, азореллу (Azorella) из семейства зонтичных, в качестве примера видов - лютик бесстебельный (Ranunculus acaulis), осоку трехраздельную (Carex trifida). Голантарктическое царство значительно уступает по числу видов всем остальным царствам. Здесь насчитывается в общей сложности немногим более 2000 видов, однако своеобразие флоры достаточно велико. Имеется десять эндемичных семейств. Все они очень малочисленные, насчитывают немного видов, а иногда только один. Эндемичных или почти эндемичных родов довольно много, большинство их характеризуется разорванным ареалом. Эндемизм на видовом уровне очень высокий - около 75%. Для Голантарктического царства характерен род антарктический, или южный, бук (Nothofagus). Представители этого рода - деревья и кустарники, близкородственные букам Северного полушария. Одни из них вечнозеленые, другие листопадные. В пределах рассматриваемого царства виды антарктического бука встречаются только в Южной Америке и Новой Зеландии. Среди примечательных представителей флоры Голантарктического царства можно назвать род подокарпус (Podocarpus) - один из древних примитивных родов хвойных, виды подокарпусов встречаются в Южной Америке и Новой Зеландии.
Учение о флорах Флорой называют совокупность видов растений, обитающих на определенной территории. Можно говорить о флоре отдельного района, области, страны или какого-либо физико-географического региона (например, флора Сибири, флора Европы, флора Омской области и т.п.). Нередко под флорой подразумевают также список растений, отмеченных на данной территории. Флоры разных территорий значительно различаются по числу слагающих их видов. Это связано, прежде всего, с размером территории. Чем она больше, тем, как правило, больше и число видов. Сравнивая приблизительно одинаковые по величине части суши по количеству видов произрастающих на них растений, выявляют флоры бедные и флоры богатые. Наиболее богаты видами флоры тропических стран, по мере удаления от экваториальной области число видов быстро уменьшается. Самой богатой является флора Юго-Восточной Азии с архипелагом Зондских островов - более 45 тыс. видов растений. На втором месте по богатству стоит флора тропической Америки (бассейн Амазонки с Бразилией) - около 40 тыс. видов. Флора Арктики - одна из самых бедных, в ней насчитывается немногим более 600 видов, флора пустыни Сахара еще беднее - около 500 видов. Богатство флоры определяется также разнообразием природных условий в пределах территории. Чем разнообразнее условия среды, тем больше возможностей для существования различных растений, тем богаче флора. Поэтому флоры горных систем, как правило, богаче равнинных флор. Так, флора Кавказа насчитывает более 6000 видов, а на обширной равнине средней полосы европейской части России встречается лишь около 2300 видов. Богатство флоры может быть обусловлено и историческими причинами. Более древние флоры, возраст которых измеряется многими миллионами лет, как правило, особенно богаты видами. Здесь могли сохраниться растения, вымершие в других областях вследствие изменения климата, оледенений и т.д. Такие древние флоры находятся, например, на Дальнем Востоке и в Западном Закавказье. Молодые флоры, сформировавшиеся относительно недавно, значительно беднее видами. Между флорами разных территорий наблюдаются существенные различия по систематическому составу. В странах умеренного климата, как правило, преобладают семейства сложноцветных, бобовых, розоцветных, злаков, осоковых и крестоцветных. В засушливых областях очень обычны различные представители маревых. Тропические флоры богаты представителями орхидных, молочайных, мареновых, бобовых, злаков. В саваннах и степях на первое место выступают злаки. Среди растений, образующих флору, можно выделить группы видов со сходными ареалами. Такие группы видов получили название географических элементов флоры. Для флоры России наиболее обычны следующие географические элементы: 1. Арктические элементы - виды, ареалы которых расположены в безлесной арктической тундре, например злак дюпонция (Dupontia fisheri). Некоторые из этих растений проникают на юг в зону хвойных лесов, где встречаются, главным образом, на болотах. В этом случае говорят о субарктических элементах, например морошка (Rubus chamaemorus) и береза карликовая (Betula nana). Нередко арктические элементы имеют также фрагменты ареалов в альпийском поясе гор Европы и Сибири. Такие виды относят к аркто-альпийским элементам, например родиола розовая (Rhodiola rosea), ива травянистая (Salix herbacea) и др. 2. Бореальные элементы - являются компонентами обширной зоны хвойных лесов (тайги), протянувшейся через всю Северную Европу и Сибирь. Характерными примерами бореальных видов являются ель сибирская (Picea obovata), сосна обыкновенная (Pinus sylvestris), линнея северная (Linnaea borealis) и др. 3. Среднеевропейские (неморальные) элементы - виды, характерные для зоны широколиственных лесов Средней Европы и европейской части России, например дуб обыкновенный (Quercus robur), копытень европейский (Asarum europaeum) и др. 4. Понтические элементы - виды, ареалы которых связаны со степной зоной Евразии, например адонис весенний (Adonis vernalis), лабазник обыкновенный (Filipendula vulgaris), степная вишня (Cerasus fruticosa). 5. Средиземноморские элементы - виды, ареалы которых охватывают присредиземноморские и причерноморские страны, например земляничник мелкоплодный (Arbutus andrachne), самшит (виды рода Buxus) и др. 6. Туранско-центральноазиатские элементы - виды, ареалы которых преимущественно ограничены пустынными и полупустынными районами Средней и Центральной Азии, например многие виды полыни (Artemisia), облепиха (Hippophae rhamnoides) и др. 7. Маньчжурские элементы - виды, ареалы которых включают зарубежную Маньчжурию и российский Дальний Восток, например маньчжурский орех (Juglans mandshurica), амурский бархат (Phellodendron amurense). При ботанико-географическом анализе флоры горных массивов учитывают распределение видов по вертикали. Флоры анализируют также с точки зрения их экологического состава. При этом выявляют доли участия в составе флоры различных экологических групп и жизненных форм растений. Близкие в экологическом отношении виды объединяют в группы, которые называются экологическими элементами флоры, например высокогорный, степной, пустынный и др. В состав флоры того или иного района могут входить растения, различные по своему происхождению. При генетическом анализе флоры все ее элементы делят на автохтонные (виды, возникшие на данной территории) и аллохтонные - виды, первоначально появившиеся за пределами территории флоры и проникшие туда в результате последующего расселения (миграции). Также выясняется возраст того или иного элемента флоры, т.е. примерное время его возникновения (для автохтонных) или проникновения на территорию данной флоры (для аллохтонных элементов). Процесс формирования флор (флорогенез) сложен, и в разных случаях он протекает по-разному. Если климат какой-либо территории резко изменяется, одна флора сменяет другую. Часть видов прежней флоры погибает, часть переселяется в другие районы, некоторые приспосабливаются к новым условиям и остаются. Вместе с тем появляется много растений из других областей, хорошо приспособленных к изменившейся природной обстановке. Если эти пришлые виды составляют основу новой флоры, такая флора носит миграционный характер. Типичной миграционной флорой является флора Арктики и большинства равнинных районов Евразии, подвергшихся в четвертичном периоде оледенению. Здесь растительный покров был полностью уничтожен, и формирование флоры шло исключительно за счет переселения растений с соседних территорий. Наряду с этим в тропических и отчасти в субтропических широтах имеются территории, не испытавшие существенных геологических и климатических изменений на протяжении сотни миллионов лет. По происхождению большинства слагающих их видов такие флоры являются автохтонными. Они считаются древними флорами, так как их современный состав сложился очень давно и с тех пор существенно не изменялся. В систематическом отношении автохтонные флоры отличаются большой целостностью. Миграционные флоры, напротив, обычно являются молодыми и разнородными в систематическом отношении. Например, флора Кергеленских островов, лежащих уединенно в Южном полушарии, включает 25 видов, относящихся к 18 родам и 11 семействам. Одна из важных особенностей любой флоры - присутствие эндемичных и реликтовых растений. Присутствие во флоре в значительном количестве эндемичных видов указывает на ее древность. Это - свидетельство того, что данная флора длительное время развивалась изолированно от остального растительного мира. Особенно богаты эндемичными видами флоры древних островов. Так, на Гавайских островах указывается 82% эндемиков, во флоре Новой Зеландии - 82%, на Мадагаскаре - 66%. Такие флоры выделяют в категорию эндемичных. Из материковых флор самой эндемичной является флора Австралии, в которой около 75% видов являются эндемиками. Количеством эндемиков определяется своеобразие, самобытность флоры. Степень эндемизма зависит от степени изолированности территории, от наличия преград, затрудняющих расселение растений и обмен видами между смежными областями, как в современную эпоху, так и в прошлом. Под реликтами подразумеваются виды, входящие в состав ныне существующей флоры, но являющиеся остатками флор минувших геологических эпох. Присутствие реликтов в какой-либо флоре также указывает на ее древность. Вместе с тем, это свидетельство того, что климат соответствующей территории сравнительно мало изменялся на протяжении всего периода существования реликтовых растений. О том, что то или иное растение относится к реликтам, судят, прежде всего, по палеоботаническим данным. Различают реликты разного возраста, сохранившиеся с тех или иных периодов геологической истории. Древнейшие реликты во флоре земного шара датируются мезозойской эрой. К таким реликтам относится, например, гинкго (Ginkgo biloba), а также секвойя (Sequoia sempervirens) и мамонтово дерево (Sequoiadendron giganteum). Это так называемые систематические реликты, единственные представители родов, семейств или даже классов, дожившие до наших дней. Более многочисленны реликты третичного периода, возраст которых значительно меньше. В третичное время, отличавшееся теплым климатом, эти растения были широко распространены по территории земного шара (в особенности в Евразии и Северной Америке). Позднее, при наступлении ледника и общем похолодании климата, теплолюбивые представители третичной флоры во многих районах погибли. Уцелели они только в отдельных убежищах (рефугиумах), где климат изменился сравнительно мало. Главнейшие рефугиумы третичной флоры Северного полушария находятся на юго-востоке Северной Америки, в Японии и Китае. На территории Северной Америки сохранились такие третичные реликты, как тюльпанное дерево (Liriodendron tulipiferum), болотный кипарис (Taxodium), некоторые магнолии (Magnolia) и многие другие растения. Очень богат третичными реликтами японо-китайский рефугиум (различные виды дуба, бука, каштана, магнолий и т. д.). Довольно много третичных реликтов и в рефугиуме на нашем Дальнем Востоке (Приморье). Среди них можно назвать амурский бархат, женьшень (Раnах ginseng), водное растение бразению (Brasenia schreberi), лотос(Nelumbo komarovii) и др. Во флоре Сибири центром концентрации третичных реликтов являются горы Южной Сибири: Приуралье, Горная Шория, Алтай, Саяны. Здесь сохранились липа (Tilia cordata и T. sibirica), копытень европейский (Asarum europaeum), щитовник мужской (Dryopteris filix-mas), бруннера сибирская (Brunnera sibirica), воронец колосистый (Actaea spicata), подмаренник душистый (Galium odoratum), двулепестник парижский (Circaea lutetiana) и др. Еще меньший возраст имеют реликты ледникового периода, или гляциальные реликты. Эти сравнительно холодостойкие растения пережили оледенение на территориях, которые не были покрыты ледником, но располагались неподалеку от него. После того, как ледник отступил, они остались на своем прежнем месте. В качестве примера таких реликтов можно назвать багульник (Ledum palustre), клюкву (Oxycoccus), бруснику (Vaccinium vitis-idaea), которые кое-где растут сейчас на болотах Среднерусской возвышенности. Следует подчеркнуть, что перечисленные растения имеют очень широкое географическое распространение, но ледниковыми реликтами они являются только на Среднерусской возвышенности. Наконец, самыми “молодыми” являются послеледниковые реликты, или реликты ксеротермического периода. В этот теплый и сухой период послеледниковья южные растения, в особенности степные, проникали далеко на север. Когда же вновь наступило похолодание климата, растения стали в массе отступать к югу. Однако кое-где они все же сохранились до настоящего времени даже далеко на севере. Таковы, например, некоторые степные растения, встречающиеся на территории Прибалтики, под Санкт-Петербургом, в ряде северных областей европейской части России, в Якутии и в других районах. При флористических исследованиях часто используется метод так называемых конкретных флор, разработанный А.И. Толмачевым. Конкретной флорой называют совокупность растений небольшой территории (на равнинах - порядка 100-500 км2), которая сравнительно однородна в природном отношении. При общем однообразии климата отдельные виды растений распределяются только в зависимости от эдафических условий и особенностей рельефа. В сходных условиях в одних и тех же местообитаниях повторяется почти полностью определенная совокупность видов. При изучении конкретной флоры выявляются и обследуются все основные местообитания, характерные для данного района, и выявляются практически все присутствующие здесь виды. В современную эпоху очень велико воздействие на флору человека и его хозяйственной деятельности. Год от года размеры территорий, где уничтожен естественный растительный покров, все возрастают. Вследствие сокращения площадей, занятых естественной растительностью, резко уменьшаются возможности произрастания многих дикорастущих видов растений, сужается круг пригодных для их жизни местообитаний. Огромное влияние на состав флоры оказывают проводимые на больших площадях распашка земель, вырубка лесов, выпас скота, массовый туризм, сбор цветов, лекарственных растений и т.п. Все эти формы деятельности человека приводят к уменьшению численности отдельных видов вплоть до их полного исчезновения. Некоторые растения оказались на грани полного уничтожения на всей площади их ареала в пределах России и занесены в “Красную книгу”. Вместе с тем, деятельность человека вызывает появление в составе флоры новых растений, совершенно несвойственных прежде той или иной местности. Это заносные, или адвентивные, виды. Они широко распространяются вдоль шоссейных и особенно железных дорог, по окраинам полей, в населенных пунктах и на других нарушенных местообитаниях, связанных с деятельностью человека. Семена подобных растений заносятся случайно и подчас издалека, даже с других континентов. Так произошло, например, с ромашкой пахучей (Chamomilla suaveolens),попавшей в свое время в Россию из Северной Америки. Это растение у нас сейчас очень широко распространилось, но встречается почти исключительно на нарушенных местообитаниях. В последние десятилетия сильно обогащается адвентивными растениями флора крупных городов. Местная флора пополняется также благодаря тому, что человек специально культивирует полезные (пищевые, декоративные) растения из других, нередко очень отдаленных районов земного шара. Большинство этих растений может существовать только в культуре, но некоторые из них дичают и внедряются в местную флору.
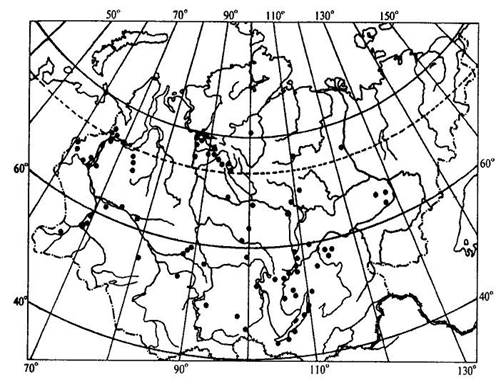
Учение об ареалах Ареал - это область географического распространения систематической единицы (вида, рода, семейства и т. д.). География растений имеет дело, прежде всего, с ареалами различных видов. Зная ареалы видов какого-либо рода, можно составить представление об ареале всего рода в целом. Так же обстоит дело и с ареалом семейства, который складывается из ареалов отдельных родов. Местонахождение - это конкретный географический пункт, где найдено или наблюдалось отдельное растение. Документом, подтверждающим нахождение в конкретном пункте определенного растения, служит, прежде всего, гербарный образец с указанием места его сбора. Пункты местонахождений отдельных растений, относящихся к изучаемому виду, могут быть нанесены на карту в виде точек. Карты с нанесенными на них точками - местонахождениями получили название точечных карт ареалов (рис. 15.14 ). Если соединить контурной линией все периферийные точки - местонахождения, то такую карту ареала называют точечно-контурной (рис. 15.15). Если распространение вида в пределах ареала не имеет существенного значения в окончательном варианте карты, территорию ареала обводят контурной линией и заштриховывают. Такая карта ареала называется контурно-штриховой (рис. 15.16).
Рис. 15.14. Пример точечной карты ареала (ареал зубровки арктической (Hierochloe arctica)). Ни один вид не занимает всю территорию своего ареала сплошь. Это связано с тем, что даже в небольшом, относительно однородном географическом районе не наблюдается полной выравненности экологических условий. Как правило, ареалы растений охватывают значительные части суши со сложной топографией и множеством разнообразных местообитаний. Каждый вид нуждается по своей экологической природе в строго определенном местообитании, таким образом, неоднородность физико-географических условий на территории ареала неизбежно порождает прерывистое распределение особей и популяций вида, следовательно, можно говорить о топографической прерывистости ареала. Однако топографическая прерывистость ареала не нарушает его географической целостности. Ареал считается сплошным при непрерывном географическом распространении вида, хотя и при наличии прерывистости топографической. Растения водоемов, солончаков, болот, песчаных почв, приуроченные к специфическим местообитаниям, нередко обнаруживают особенно значительную прерывистость своего топографического распределения. Промежуточные пространства с иными местообитаниями заполняются популяциями других видов в соответствии с их экологической приспособленностью. В результате этого один и тот же географический район может входить в ареалы многих видов, т. е. ареалы разных растений обычно налагаются друг на друга в тех или иных частях. Например, в Западной Сибири произрастают такие древесные породы, как сибирский кедр, пихта, ель, сосна. Эти виды находятся здесь в пределах своего ареала, но в то же время каждый занимает свое местообитание: сосна на песчаных террасах рек, ель в поймах, на богатых наносных почвах, кедр и пихта на суглинках ближе к водоразделам. Ареалы этих видов налагаются только в определенной части равнины, тогда как размеры и общие очертания ареалов значительно расходятся.
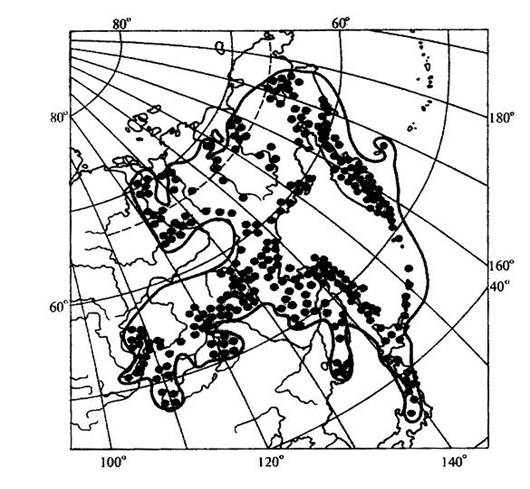
Рис. 15.15. Пример точечно-контурной карты ареала ( ареал кедрового стланика (Pinus pumila)). Рассматривая географическое распространение растений, мы можем иногда встретиться с таким фактом, когда промежуток между отдельными местонахождениями вида охватывает значительную часть суши, которая не может быть включена в единый ареал данного вида, так как здесь имеет место географическая прерывистость. В этом случае говорят о разрыве ареала, или его дизъюнкции. Дизъюнкция представляет собой довольно распространенное явление, особенно в родах и семействах. Таковы ареалы родов орех (Juglans), к которому относятся о. грецкий (J. regia), о. маньчжурский (J. mandshurica) и ряд американских видов; саговник (Cycas) и многих других. Разорванные (дизъюнктивные) ареалы обычно являются наследием геологического прошлого, т. е. представляют остатки некогда сплошных ареалов. Подтверждением этому являются ископаемые остатки растений, находимые на территориях современных разрывов ареалов. Например, в третичных отложениях северной Евразии, в том числе и Сибири, широко известны остатки орехов, родственных грецкому ореху, что свидетельствует о былом обширном ареале этого рода, имеющем в настоящее время дизъюнктивный характер.
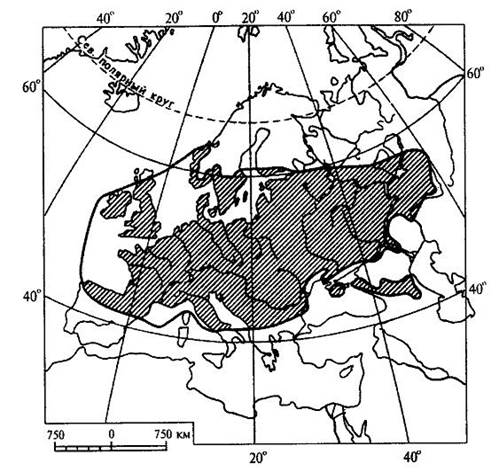
Рис. 15.16. Пример контурно-штриховой карты ареала (ареал дуба обыкновенного (Quercus robur)). Ареал образуется в результате расселения вида и последовательного завоевания им определенного географического пространства и является результатом взаимодействия биологических особенностей вида с физико-географическими условиями в их историческом развитии. Следовательно, к основным факторам формирования ареалов можно отнести следующие: 1. Биологические особенности растений: а) естественные способы распространения; б) экологическая амплитуда. 2. Современные физико-географические условия, в которых происходит расселение вида. 3. Возраст вида. 4. Изменения физико-географических условий материков в геоисторическом масштабе. 5. Деятельность человека. В зависимости от тех причин, которые обусловливают границы ареалов, в географии растений различают следующие типы границ: климатические, непереходимые, почвенно-грунтовые, биотические, исторические. 1. Климатические границы возникают как следствие приспособленности растений к режиму тепла и влаги той природной зоны, где каждый данный вид существует в оптимальных условиях. У большинства растений Северного полушария, особенно древесных, полярная граница является климатической (ель, лиственница и др). Зависимость границ ареалов от климата проявляется в том, что во многих случаях эти границы идут по определенным параллелям или повторяют изолинии каких-либо климатических показателей. Наглядным примером являются северная и южная границы ареала семейства пальм, которые проходят по северному и южному тропикам. 2. Непереходимые границы возникают за счет физических преград, встающих на пути распространения растений. Сюда относятся обширные водные пространства - океаны, моря, высокие горные хребты, особенно в случае их широтного простирания. Эту же роль играют большие площади открытых ландшафтов пустынь, степей, тундр - для лесных растений и значительные лесные пространства - для растений степей и пустынь. 3. Почвенно-грунтовые, или эдафические, границы образуются при исчезновении необходимых местообитаний в связи с резким изменением химизма или физических свойств почв. Так, сильная засоленность северной половины Крымского полуострова является препятствием для расселения сюда многих степных видов, распространенных на юге Украины. Географически сплошная вечная мерзлота на северо-востоке Сибири является причиной образования здесь границ ареалов ряда древесных пород, обычных в остальной Сибири, таких как ель, сосна, сибирский кедр. 4. Биотические границы иногда называют еще конкурентными, так как связывают их с невозможностью внедрения вида в какую-либо местность потому, что необходимые для него местообитания уже раньше успели занять популяции другого вида со сходными экологическими требованиями. Именно это обстоятельство нередко ограничивает роль естественных факторов географического распространения растений. Виды, обосновавшиеся ранее в данной местности, “не пускают” пришельцев, не дают им размножиться и продвинуться за какую-то достигнутую ими границу. Биотическая граница может возникнуть также в том случае, если вид в процессе своего расселения достигает района, где по какой-либо причине отсутствует необходимое для него насекомое - опылитель. Не имея возможности обеспечить себя потомством, вид не перейдет через границу ареала этого насекомого. 5. Исторические границы, в отличие от предыдущих типов, обусловлены не современными причинами, а какими-нибудь событиями геологического прошлого, такими, как фазы горообразования, изменения конфигурации и связей материков, изменения климата. К историческим границам можно отнести также те случаи, когда вид в современную эпоху активно расселяется и не достиг еще потенциально возможных для него пределов. К такому типу можно отнести западную границу ареала сибирского кедра (рис. 15.17 ), который в своем расселении из Западной Сибири перевалил через Урал и Приуралье и еще не успел проникнуть дальше на запад.
Рис. 15.17. Ареал кедра сибирского (Pinus sibirica). Таким образом, у одного и того же вида разные границы его ареала могут иметь разное происхождение и относиться к различным типам. Так, северная, южная и отчасти восточная границы ареала сибирского кедра являются климатическими, северо-восточная - эдафической, а западная - исторической. Ареал в своем развитии проходит определенные стадии. Первоначально, в момент своего возникновения, вид часто имеет незначительный по площади ареал (первичный ареал). Затем, если условия среды благоприятны, происходит расселение вида, и ареал постепенно расширяется. В конце концов, вид достигает своих пределов распространения, ареал перестает существенно увеличиваться по площади, и его границы более или менее стабилизируются. Когда вид расселяется, его ареал находится в стадии прогресса. Однако ареалы могут претерпевать и обратные, регрессивные изменения, сокращаясь по площади. Это происходит вследствие изменения окружающей среды в неблагоприятном для вида направлении (например, похолодание или иссушение климата). В конечном счете, от некогда обширного ареала может остаться лишь очень небольшая его часть. Такие ареалы называют реликтовыми. Примерами растений, имеющих реликтовые ареалы, могут быть гинкго двулопастный (Ginkgo biloba), мамонтово дерево, или секвойядендрон гигантский (Sequoiadendron giganteum), болотный кипарис (Taxodium distichum) и др. В зависимости от формы ареалы делят на две основные группы: 1) сплошные и 2) несплошные. Наиболее распространенными формами среди сплошных ареалов являются: 1. Опоясывающие - вытянутые вдоль всей суши земного шара по широте, это ареалы целых семейств или родов (семейства березовых, сосновых, пальм и др.). 2. Циркумполярные - охватывающие полярную окраину северной суши полукольцом или почти кольцом. 3. Овальные - вытянутые в меридиональном направлении или чаще по широте на ограниченном отрезке того или иного климатического пояса, наиболее распространенная форма ареалов видов и родов, но встречается и у семейств. 4. Лучистые и бахромчатые - обычно неправильной формы с неравномерными, часто многочисленными выступами в разных направлениях, это ареалы активно расселяющихся видов. К несплошным ареалам относятся: 1. Дизъюнктивные, или разорванные, - регрессивные ареалы, распадающиеся на два или несколько изолированных фрагмента (семейство саговниковых, роды орех, каштан и др.); дизъюнктивный ареал может возникнуть и вследствие случайного заноса семян, например перелетными птицами, в пригодные для произрастания вида места, находящиеся на значительном расстоянии от границ его основного ареала. 2. Продырявленные - встречаются у вымирающих папоротников и некоторых видов цветковых растений. 3. Точечные ареалы - слагаются из многочисленных, сильно разъединенных участков, характерны для водных, сорных и рудеральных (мусорных) растений. 4. Ленточные ареалы - представляют собой фрагменты, оторванные от основного, сплошного, ареала и разъединенные большими пространствами, непригодными для обитания данного вида; чаще всего это ареалы древесных видов и сопутствующих им трав и кустарников, проникающих в тундры, пустыни, саванны вдоль рек, около которых они и протягиваются узкими лентами. Размеры ареалов могут колебаться в широком диапазоне. Особенно значительные площади занимают опоясывающие ареалы некоторых семейств, некоторые роды (береза, сосна) также имеют очень обширные сплошные ареалы. Но ареал вида обычно ограничен какой-либо климатической зоной, охватывая лишь отдельную ее часть. Виды, встречающиеся на всех континентах, называются космополитами. К ним относятся некоторые водные растения, например элодея канадская (Elodea canadensis), ряска малая (Lemna minor), тростник обыкновенный (Phragmites australis) и др. Обитатели водной среды никогда не испытывают недостатка во влаге и мало зависят от климата. Кроме того, эти растения способны быстро расселяться по территории, так как их семена, плоды, вегетативные органы легко разносятся водой или птицами. Обширные ареалы имеют некоторые сорные растения, произрастающие на нарушенных местообитаниях, где ослаблена конкуренция между растениями, и сопутствующие человеку на всех материках, например пастушья сумка (Capsella bursa-pastoris), крапива двудомная (Urtica dioica) и ряд других. Отдельные местонахождения этих растений, как правило, настолько удалены друг от друга, что соединять их в один сплошной ареал было бы неправильно. Подобные ареалы обычно показывают на картах рассеянными точками, и ареал называют точечным. Если растение распространено в пределах примерно половины суши, то его относят к полукосмополитам. Последние наиболее часто встречаются среди споровых растений, например папоротник орляк (Pteridium aquilinum), некоторые мхи. Широко распространенные виды называют эврихорами, а виды, ограниченные в распространении, - стенохорами. Все космополиты - эврихоры, но не все эврихоры являются космополитами, так как некоторые из них встречаются повсеместно лишь на одном из континентов. В природе количественно преобладают мезохоры и стенохоры. К этим двум группам относится около 70% всех видов растений. Среди стенохоров имеются виды, ареал которых ограничен только каким-либо определенным регионом, и вне его они нигде больше не встречаются. Подобные растения называют эндемичными, или эндемиками (эндемами). Можно говорить, например, об эндемиках Кавказа, Крыма, Средней Азии, Алтая и т.д. Эндемичными могут быть не только виды, но и другие таксоны (роды, семейства и т.д.). Одни районы земного шара более богаты эндемиками, другие - менее, в третьих - эндемики отсутствуют. Особенно велика доля эндемичных растений во флоре древних морских островов. Так, во флоре Новой Зеландии насчитывается около 75% эндемичных видов. Древние острова особенно богаты эндемиками потому, что их растительный мир на протяжении очень длительного времени был полностью изолирован, и никакого обмена растениями с другими территориями не происходило. В этих условиях на островах в ходе эволюции возникли новые виды. Наряду с этим здесь могли сохраниться и некоторые растения, вымершие на остальной территории земного шара. Сравнительно велик процент эндемичных растений также в высоких поясах древних горных систем (субальпийском и альпийском). Причина этого явления - отсутствие обмена растениями с окружающими равнинными территориями. Высокогорья подобны островам в океане, флора их изолирована. В горных системах более молодого возраста эндемиков значительно меньше. Различают два типа эндемизма - палеоэндемизм (реликтовый эндемизм) и неоэндемизм. В первом случае речь идет о вымирающих таксонах, которые прежде были широко распространены, но сейчас встречаются лишь на очень ограниченных территориях. Например, гинкго двулопастный (Ginkgo biloba) - единственный современный представитель целого класса голосеменных растений, который в мезозойскую эру был широко распространен по всей Евразии и Северной Америке, - растет в диком состоянии на очень небольшой территории в горах Восточного Китая. Мамонтово дерево (Sequoiadendron giganteum) встречается только в горах северной Калифорнии, тогда как в третичном периоде было распространено по всему Северному полушарию. Такие виды можно назвать реликтовыми эндемиками. В случае неоэндемизма речь идет, напротив, о молодых видах. Их ареал ограничен, главным образом, по той причине, что они еще не успели достаточно широко распространиться по земной поверхности. Примером неоэндемиков могут быть некоторые “молодые” виды первоцвета (Primula), колокольчика (Campanula), крупки (Draba), встречающиеся на Кавказе. Часто встречается явление викаризма. Викарирующими, или викарными (замещающими), называют виды и другие таксоны, которые мало различаются по морфологическим признакам и близко родственны между собой, но территориально разобщены. Эти виды как бы замещают друг друга на разной территории. В качестве примера можно назвать два вида ели, произрастающих в России, - ель европейскую (Picea abies) и ель сибирскую (P. obovata). По морфологическим признакам они довольно похожи, но географическое распространение их различно. Ареал первого вида расположен в европейской части России, второго – включает северо-восток европейской части, Сибирь и Дальний Восток. Хорошим примером викаризма служат также виды лиственницы (Larix), которые в разных частях Северного полушария образовали почти непрерывный ряд форм, замещающих одна другую.
Геоботаника Геоботаника – раздел ботанической географии, изучающий сложение и распространение по поверхности Земли растительного покрова. Разделами геоботаники являются: 1) фитоценология – учение о растительных сообществах (фитоценозах); 2) география растительности – учение о закономерностях географического распределения растительных сообществ и сочетании их в пространстве. Основными объектами исследований в геоботанике являются: 1) растительное сообщество, или фитоценоз, и 2) растительный покров, или растительность. Под растительным сообществом (фитоценозом) понимают конкретную группировку растений в пределах одного местообитания, характеризующуюся определенным видовым составом, строением и определенной системой взаимоотношений растений друг с другом и со средой. Любая более или менее крупная территория расчленяется на значительное число разнообразных местообитаний, в каждом из которых образуется особый фитоценоз. Всю совокупность фитоценозов той или иной территории называют ее растительным покровом, или растительностью. Так, например, совокупность фитоценозов на территории Западно-Сибирской равнины образует растительный покров Западно-Сибирской равнины. Совокупность фитоценозов всей земной суши образует растительный покров Земли, для обозначения которого используется также термин «фитосфера».
Фитоценология Фитоценозы являются частью более сложных природных систем – биогеоценозов, совокупность которых образует биосферу Земли. Биогеоценоз - система, состоящая из косной (неживой) среды – экотопа и живых организмов, образующих биоценоз. В состав биоценоза входят растительность (фитоценоз), животное население (зооценоз) и совокупность микроорганизмов (микробоценоз). Ведущую роль в биогеоценозе играет фитоценоз, обладающий огромной биомассой. Фитоценоз сильно влияет на атмосферу, почву, определяет видовой состав животных, микроорганизмов, а также многие особенности обмена веществ и энергии в биогеоценозе. Границы биогеоценоза в горизонтальном направлении определяются границами фитоценоза, свойственного ему, в вертикальном – высотой надземных органов фотосинтезирующих растений и глубиной проникновения их подземных органов. Фитоценозы представляют собой результат длительного подбора видов растений, которые в процессе взаимодействия между собой и другими компонентами биоценоза приспособились к определенным экологическим условиям. В результате совместного произрастания в фитоценозе устанавливаются определенные взаимоотношения между видами. Принято различать три основные формы влияния растений друг на друга: прямые (контактные), трансабиотические, трансбиотические. Контактные отношения проявляются в паразитизме, симбиозе, в механическом воздействии растений друг на друга, в срастании корней и т.п. Трансабиотические взаимоотношения – это влияние одних растений на другие посредством изменения окружающей среды. Сюда относится конкуренция за свет, влагу, питательные вещества, а также выделение растениями продуктов жизнедеятельности в окружающую среду и др. Трансбиотические взаимоотношения – это влияние одних растений на другие через посредство каких-то иных организмов. Обычно в фитоценозах одновременно проявляются различные формы взаимного влияния растений. При формировании растительных сообществ большую роль играют конкурентные взаимоотношения как между особями одного вида, так и между особями разных видов. Несмотря на наличие межвидовой конкуренции, практически в каждом естественном фитоценозе возможно сосуществование нескольких или многих видов растений. Возможность эта обусловлена тем, что каждый вид в фитоценозе занимает свое особое место, которое называют экологической нишей. Структура фитоценозов.Важнейшим признаком фитоценоза является его видовой (флористический) состав. Число видов растений в сообществе называют его видовым богатством. Оно зависит от ряда факторов. Большую роль играют общие физико-географические и исторические условия, от которых зависит видовое богатство флоры каждого конкретного района. Так, например, видовая насыщенность фитоценозов влажных тропических лесов, формирующихся в условиях исключительно богатой тропической флоры, оценивается сотнями видов высших растений, а видовая насыщенность сибирских таежных лесов, формирующихся на фоне бедной бореальной флоры, варьирует, как правило, в пределах 15-30 видов. Флористическое разнообразие фитоценозов зависит также от условий местообитаний: чем они благоприятнее, тем сложнее видовой состав, и, наоборот, в неблагоприятных местообитаниях формируются флористически простые фитоценозы. Каждый вид в фитоценозе представлен, как правило, более или менее значительным числом особей. Совокупность особей одного вида, входящих в состав определенного растительного сообщества, называется ценопопуляцией (ценотической популяцией). Каждая ценопопуляция занимает в фитоценозе определенное положение, т.е. играет определенную роль, которая зависит от следующих признаков ценопопуляции: количественного участия, возрастного состава, фенологического состояния, жизненного состояния, эколого-биологических особенностей. Количественное участие видов в растительных сообществах, обычно называемое в геоботанике обилием, выражается различными показателями: численностью особей, проективным покрытием поверхности почвы их надземными частями, массой, объемом. В большинстве фитоценозов есть виды, которые по численности особей преобладают над другими. Такие виды называют доминантами. По количеству доминантов различают фитоценозы моно- и полидоминантные (имеется лишь один господствующий вид или же их несколько). Примером монодоминантных сообществ могут быть многие леса таежной зоны, где в древесном ярусе господствует какой-либо один вид (ель, сосна и др.). Дубравы, напротив, обычно полидоминантны. В травяных сообществах (луга, степи) нередко происходит смена доминантов по годам в зависимости от изменения погодных условий. Очень трудно выделить доминирующие виды в дождевых тропических лесах, в некоторых типах лугов и ряде других сообществ. Особи, входящие в состав ценопопуляции, как правило, находятся на разных стадиях онтогенеза, или в разных возрастных состояниях, начиная от еще не проросших и прорастающих семян и кончая старыми отмирающими экземплярами. Распределение особей ценопопуляции по возрастным состояниям называют возрастным спектром ценопопуляции. В зависимости от соотношения возрастных групп различают три типа ценопопуляций: инвазионный, нормальный и регрессивный. Инвазионные популяции состоят исключительно или в основном из молодых особей, еще не достигших половозрелого состояния и не дающих генеративного потомства. Нормальные ценопопуляции характеризуются гармоничным сочетанием молодых, половозрелых и стареющих особей. В регрессивных ценопопуляциях преобладают стареющие особи при полном или почти полном отсутствии молодых возрастных групп. Принадлежность ценопопуляции к определенному возрастному типу характеризует ее позицию и жизнеспособность в фитоценозе. Анализ возрастного состава ценопопуляций позволяет выяснять состояние отдельных ценопопуляций и фитоценоза в целом, прогнозировать направление их дальнейшего развития, помогает разрабатывать режим рационального использования фитоценозов, решать задачи их оптимизации и охраны. Каждый растительный организм в течение вегетационного периода проходит ряд стадий сезонного роста и развития, начиная от прорастания генеративных зачатков или от возобновления роста перезимовавших почек и побегов до отмирания растения или подготовки к новому зимнему периоду. Эти стадии развития растений, адаптированные к сезонной динамике факторов среды, называют фенологическими фазами, или фенофазами. Результаты фенологических исследований отражают в виде так называемых фенологических спектров, на которых показывается последовательность и длительность прохождения фенологических фаз особи, ценопопуляции или фитоценоза в целом. Характер фенологических спектров зависит, с одной стороны, от видовой специфичности растений и, с другой стороны, от особенностей сезонных изменений экологических условий. Поэтому анализ фенологических спектров ценопопуляций позволяет выявлять экологию видов, фенологическое состояние ценопопуляций в каждый конкретный момент времени, их годовой цикл развития, изменение роли ценопопуляций в течение вегетационного периода, а также фазы сезонного развития всего фитоценоза. Жизненность ценопопуляции определяется жизненным состоянием образующих ее особей, а под жизненностью особей понимают их свойства, проявляющиеся в мощности особей и различных перспективах их развития. Жизненность особей характеризуется такими показателями, как возрастное состояние, диаметр, высота и степень разветвленности побегов, размер листовых пластинок, число генеративных побегов и т.д. Жизненность ценопопуляций изменяется во времени и при переходе от одного фитоценоза к другому. Поэтому изучение жизненного состояния ценопопуляций помогает определить положение и роль каждой ценопопуляции в конкретном фитоценозе. Каждая ценопопуляция в фитоценозе играет определенную роль, влияя на формирование его внутренней среды (фитосреды) и на другие ценопопуляции. Роль ценопопуляции зависит не только от ее количественного участия, но и от эколого-биологических особенностей вида. Обычно в составе фитоценоза различают три группы видов, обладающих сходным средообразующим воздействием и сходной степенью влияния на другие ценопопуляции фитоценоза. 1. Эдификаторы – ценопопуляции, слагающие основу фитоценоза, играющие главную роль в формировании фитосреды, оказывающие большое влияние на другие ценопопуляции. Они являются доминантами и образуют основную фитомассу в фитоценозе. Чаще всего это растения верхних ярусов, например лиственница в лиственничном лесу, ель в еловом лесу. Иногда эдификаторами могут быть растения нижнего яруса, например сфагнум на верховом болоте. Эдификаторами являются устойчиво доминирующие виды, оказывающие большое влияние на формирование фитосреды. 2. Ассектаторы – ценопопуляции, характерные для фитоценоза, постоянно присутствующие в нем, но не обильные и потому не имеющие большого фитоценотического значения. 3. Адвентивные растения – ценопопуляции случайные, не свойственные данному фитоценозу, быстро исчезающие из него, например полевые сорняки на естественных лугах, крапива в водораздельных лесах и т.д. Фитоценозы расчленяются на отдельные морфологические части как в вертикальном, так и в горизонтальном направлении, в соответствии с этим различают вертикальную и горизонтальную структуру фитоценозов. Вертикальная структура фитоценозов обусловливается тем, что произрастающие в нем растения имеют неодинаковую высоту, а их корневые системы проникают в почву на разную глубину. В результате фитоценоз расчленяется в вертикальном направлении на отдельные более или менее отграниченные друг от друга слои – ярусы, что приводит к более полному использованию растениями ресурсов местообитания. Наиболее четко ярусность выражена в лесном фитоценозе. Число ярусов в разных типах леса различно. Например, сосняк сфагновый может иметь всего два яруса: в первом – сосна, во втором – сфагнум. В дубраве можно выделить иногда до семи ярусов. Обычно в лесу выделяют: 1) древостой, или древесный ярус; 2) подлесок, или кустарниковый ярус; 3) травяно-кустарничковый ярус (травы, кустарнички и полукустарнички); 4) мохово-лишайниковый ярус. В фитоценозах есть также внеярусные растения, к ним относятся лианы и эпифиты. В некоторых типах фитоценозов вертикальная структура бывает выражена слабо. Так, например, во влажных тропических лесах, где плотность растительного населения особенно велика, вертикальная структура приобретает непрерывный характер и ее можно расчленить на ярусы лишь условно. Нечетко выделяется ярусность во многих травянистых фитоценозах, в которых на непрерывность вертикальной структуры в пространстве накладывается ее изменяемость во времени. Подземные органы различных растений располагаются в почве на разной глубине. Здесь тоже можно различать ярусы. Подземная ярусность хорошо выражена в лесных фитоценозах. Горизонтальная структура фитоценозов определяется характером распределения растений по их площади. В естественных фитоценозах, как правило, ценопопуляции распределяются неравномерно, при этом в одних точках особи образуют скопления (группы), между которыми особи данной ценопопуляции не встречаются совсем или встречаются значительно реже. Неравномерность распределения растений в фитоценозах обусловлена целым рядом причин: 1) особенностями размножения растений и формой их роста; 2) неоднородностью экотопа в разных точках, в которых особи находят неодинаковые условия существования; 3) взаимоотношениями растений друг с другом; 4) деятельностью животных, а также 5) разнообразными влияниями человека. В зависимости от типа распределения доминантных ценопопуляций возникает два типа горизонтальной структуры – диффузный и мозаичный. Диффузная структура характеризуется более или менее однородным (гомогенным) горизонтальным строением. Мозаичная структура характеризуется явно неоднородным (пятнистым) распределением доминантных ценопопуляций, в результате чего в фитоценозе выделяются мелкие участки, отличающиеся друг от друга по составу и строению. Наличие мозаичности имеет большое значение в функционировании фитоценозов, так как в мозаичных ценозах усложняется эколого-биологический состав ценопопуляций, полнее используются ресурсы местообитания, повышается устойчивость фитоценоза. Динамика фитоценозов. Каждый фитоценоз представляет собой не застывшее явление природы, а систему, находящуюся в состоянии постоянной динамики. Изменчивость фитоценоза во времени – это одна из наиболее характерных его особенностей. Фитоценозы претерпевают разнообразные изменения, которые можно разделить на модификации и смены. К модификациям относят обратимые изменения фитоценозов. Это сравнительно непродолжительные и неглубокие, главным образом, количественные изменения признаков фитоценозов. Вызываются они обратимыми изменениями экологических условий и являются следствием приспособительных реакций растений на изменения этих условий. При восстановлении экологических условий в исходное состояние, возвращаются в исходное состояние и все обратимые признаки фитоценозов. В зависимости от продолжительности модификаций различают суточную, сезонную и разногодичную изменчивость. Суточная изменчивость обусловлена сменой дня и ночи, с которой сопряжено изменение различных факторов местообитания. Прежде всего, изменяются условия освещения, а вместе с ними изменяются температура, влажность, воздействие животных и другие факторы. Растения реагируют на суточные изменения экологических условий изменением интенсивности процессов жизнедеятельности. Кроме того, у ряда видов растений существуют суточные ритмы в цветении, в расположении листьев или соцветий. Все это ведет к незначительным изменениям во внешнем облике и структуре фитоценозов, в то время как видовой состав в течение суток остается неизменным. Сезонная изменчивость фитоценозов, так же как и суточная, характеризуется строгой периодичностью. Она обусловлена двумя причинами – сезонными изменениями экологических условий и особенностями сезонного роста и развития растений. Флористический состав является одним из наиболее стабильных признаков, так как он практически не зависит от фенологического развития растений. Резко меняются по сезонам года состав вегетирующих и покоящихся ценопопуляций, возрастной состав (особенно у ценопопуляций однолетних растений), количественные соотношения ценопопуляций. К изменчивым признакам в сезонной динамике фитоценозов относится также аспект, под которым понимают внешний вид фитоценоза, его физиономичность в каждое конкретное время года. Смена аспектов фитоценозов определяется сезонной динамикой их структуры и сменой фенологического состояния ценопопуляций. Смену аспектов переживают все фитоценозы, но не в одинаковой степени. Для травянистых фитоценозов, в особенности для степных и пустынных с большим участием эфемерных растений, характерна резкая смена аспектов, а для вечнозеленых хвойных лесов – слабая, визуально трудно различимая смена аспектов. К разногодичным изменениям, называемым также флюктуациями, относят более продолжительные, по сравнению с сезонными, модификации фитоценозов. Это такие изменения, которые осуществляются в течение нескольких соседних лет и для которых временным масштабом измерения является один год. Флюктуации, также как и сезонные изменения, являются обратимыми и не ведут к смене одного фитоценоза другим. Однако, в отличие от сезонной изменчивости, флюктуации не отличаются строгой периодичностью, а совершаются то в одну, то в другую сторону от какого-то среднего состояния фитоценоза. Флюктуациям подвержены почти все признаки фитоценозов, но не в одинаковой степени. Наиболее устойчив флористический состав фитоценоза, достаточно сильно могут флюктуировать признаки структуры, продуктивность, характер фитосреды и т.д. Травянистые фитоценозы подвержены более резким разногодичным изменениям, нежели лесные. Наименее подвержены флюктуациям фитоценозы, образованные вечнозелеными растениями. Флюктуации могут быть вызваны различными причинами. Главнейшую роль играют внешние причины, прежде всего циклические изменения климатических и гидрологических условий. Так, во влажные годы в одном и том же фитоценозе сильнее разрастаются более влаголюбивые виды, а в засушливые, наоборот, более засухоустойчивые. Для фитоценозов пойменных лугов большое значение имеет продолжительность затопления весной. Если пойма остается под водой более длительный период, чем обычно, то в травостое большую роль начинают играть гигрофиты и даже болотные виды. Флюктуации фитоценозов бывают настолько резкими, что создается впечатление смены одного фитоценоза другим. Однако, в отличие от смен, флюктуации всегда завершаются через сравнительно непродолжительное время возвратом к исходному состоянию. К сменам относят необратимые изменения фитоценозов, которые ведут к замещению одного фитоценоза другим. В отличие от модификаций, смены носят более глубокий характер, так как затрагивают не только изменение количественных соотношений ценопопуляций, но всегда сопровождаются существенными изменениями флористического состава фитоценозов. Способность к сменам – одно из важнейших свойств фитоценозов, обусловленное двумя причинами: во-первых, тем, что фитоценозы представляют собой открытые системы, испытывающие различные воздействия внеш
Поиск по сайту: |