|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
ВОПРОС № 6. КАТАБОЛИЗМ ПУРИНОВЫХ НУКЛЕОТИДОВ
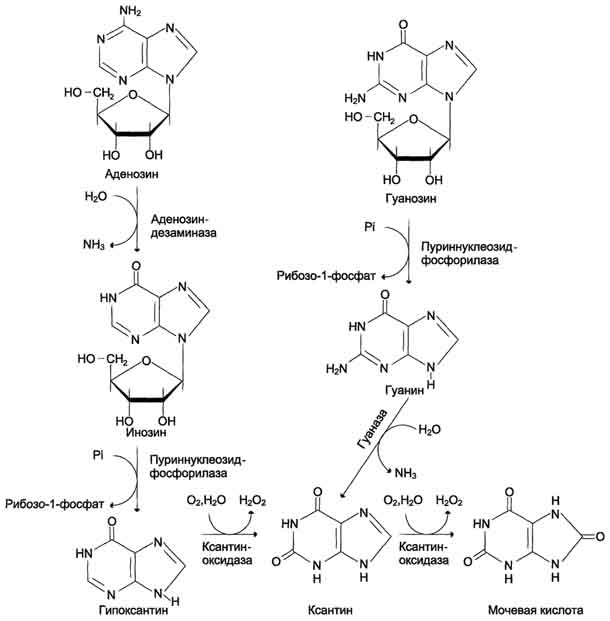
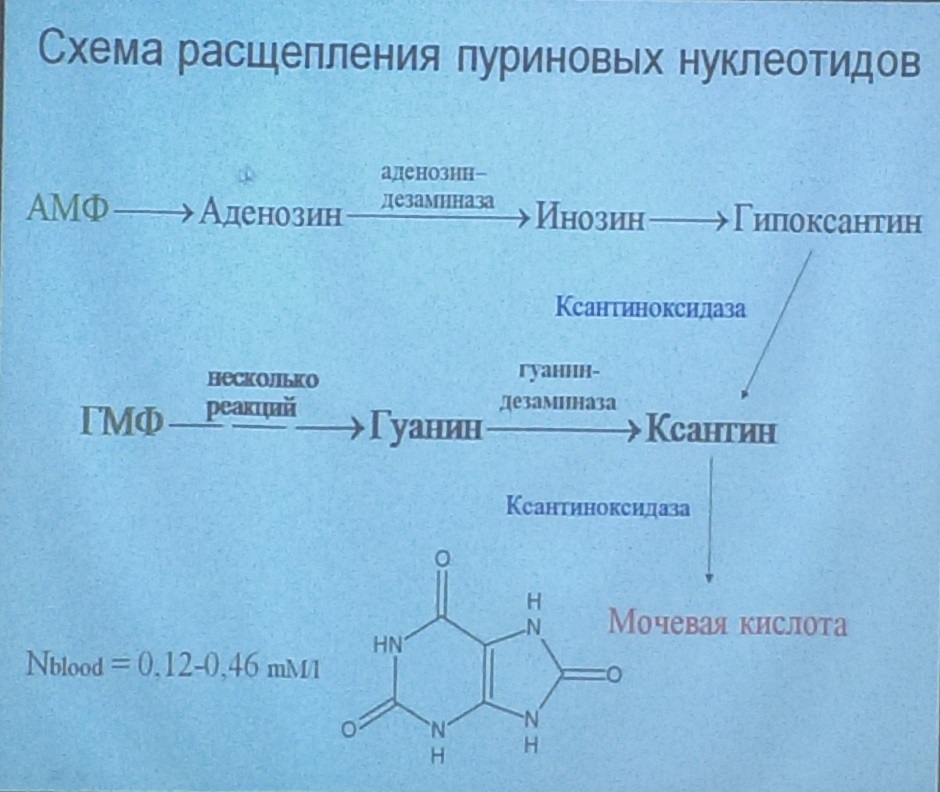
У человека основной продукт катаболизма пуриновых нуклеотидов - мочевая кислота. Её образование идёт путём гидролитического отщепления фосфатного остатка от нуклеотидов с помощью нуклеотидаз или фосфатаз, фосфоролиза N-гликозидной связи нуклеозидов пуриннуклеозидфосфорилазой, последующего дезами-нирования и окисления азотистых оснований (рис. 10-9). Распад пуриновых нуклеотидов начинается с отщепления фосфатной группировки под действием 5’-нуклеотидазы. Из аденилаты таким путем образуется аденозим, который , дезаминируясь превращается в ионозин. Ионозин подвергается гидролизу , что приводит к образованию пуринового основания гипокстантина и D-рибозы. Гипоксантин окисляется до ксантина и далее до мочевой кислоты под действием ксантинооксидазы, сложного флавинзависимого фермента в простетической группе которого содержится один атом молибдена и 4 железосерных центра. Акцептором водорода в этой реакции служит молекулярный кислород. Конечным продуктом распада ГМФ также является урат. ГМФ сначала гидролизуется с образованием нуклеозида гуанозина, который расщепляясь превращается в свободный гуанин. Гуанин претерпевает гидролитическое расщепление с образоанием ксантина, превращающегося затем в урат под действием ксантиноксидазы От АМФ и аденозина аминогруппа удаляется гидролитически аденозиндезаминазой с образованием ИМФ или инозина. ИМФ и ГМФ превращаются в соответствующие нуклеозиды: инозин и гуанозин под действием 5´-нуклеотидазы. Пуриннуклеозидфосфорилаза катализирует расщепление N-гликозидной связи в инозине и гуанозине с образованием рибозо-1-фосфата и азотистых оснований: гуанина и гипоксантина. Гуанин дезаминируется и превращается в ксантин, а гипоксантин окисляется в ксантин с помощью ксантиноксидазы, которая катализирует и дальнейшее окисление ксантина в мочевую кислоту. Ксантиноксидаза - аэробная оксидоредуктаза, простетическая группа которой включает ион молибдена, железа (Fe3+) и FAD. Подобно другим оксидазам, она окисляет пурины молекулярным кислородом с образованием пероксида водорода. В значительных количествах фермент обнаруживается только в печени и кишечнике. Мочевая кислота удаляется из организма главным образом с мочой и немного через кишечник с фекалиями. У всех млекопитающих, кроме приматов и человека, имеется фермент уриказа, расщепляющий мочевую кислоту с образованием аллантоина, хорошо растворимого в воде (рис. 10-10). Амфибии, птицы и рептилии, подобно человеку, лишены уриказы и экскретируют мочевую кислоту и гуанин в качестве конечных продуктов обмена.
Рис. 10-9. Катаболизм пуриновых нуклеотидов до мочевой кислоты. Мочевая кислота является слабой кислотой. Содержание недиссоциированной формы и солей (уратов) зависит от рН раствора. При физиологических значениях рН у мочевой кислоты может диссоциировать только один протон из трёх (рК = 5,8), поэтому в биологических жидкостях присутствует как недиссоциированная кислота в комплексе с белками, так и её натриевая соль. В сыворотке крови в норме содержание мочевой кислоты составляет 0,15-0,47 ммоль/л или 3-7 мг/дл. Ежесуточно из организма выводится от 0,4 до 0,6 г мочевой кислоты и уратов.
ВОПРОС № 7. БИОСИНТЕЗ ДЕЗОКСИРИБОНУКЛЕОТИДОВ Синтез дезоксирибонуклеотидов идёт с заметной скоростью только в тех клетках, которые вступают в S-фазу клеточного цикла и готовятся к синтезу ДНК и делению. В покоящихся клетках дезоксинуклеотиды практически отсутствуют. Все дезоксинуклеотиды, кроме тимидиловых, образуются из рибонуклеотидов путём прямого восстановления ОН-группы у второго углеродного атома рибозы в составе рибонуклеозиддифосфатов до дезоксирибозы. Тимидиловые нук-леотиды синтезируются из dУМФ особым путём с участием N5,N10-метилен-Н4-фолата.
Поиск по сайту: |