|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Избыточное поступление таурина 3.2.2.1 Однократное парентеральное введение
Однократное внутрибрюшинное введение Таи в дозе 650 мг/кг вызывает увеличение концентрации этого соединения в печени и плазме крови в среднем в 2-3 раза. В плазме крови на этом фоне увеличилось содержание цистеата, а уровни метионина, аланина, серина, АРУЦ и ААК снизились на 50% (рис.11). В печени содержание цистеина практически не изменилось, а метионина и цистеата снизилось вдвое. Так же, как и в плазме, в печени уменьшилось количество серина и, особенно — аланина, а треонина и глицина выросло (рис.11). При этом возникли высоко достоверные отрицательные связи (р<0,02, г = —0,85 — 0,90) в содержании гликогенных аминокислот и АРУЦ печени и плазмы крови. С позиций мембранотропных эффектов Таи возникшая метаболическая ситуация объясняется, прежде всего, активной экстракцией перечисленных аминокислот из кровеносного русла тканями, т.е. активацией в них метаболических процессов [202, 203]. Мембранстабилизирующее антиоксидантное действие Таи подтверждено нами в опытах in vitro, где Таи в концентрациях, соответствующих его уровню в тканях (10'4— 10'6М) (табл.12), проявлял выраженную антирадикальную активность, подавляя на 30-40% люминесценцию спиртового раствора олеата, вызванную Fe2+. В гомогенатах печени опытных животных была достоверно выше на 12% суммарная антиокислительная активность. Приблизительно на столько же увеличилось в печени содержание общих фосфолипидов и их фракций — фосфатидилхолина и сфингомиелина, уровень этаноламина снизился, а фосфоэтаноламина увеличился (рис.12). Ранее было продемонстрировано гиполипидемическое действие таурина [69]. Увеличение синтеза фосфолипидов можно также рассматривать не только как один из аспектов мембраностабилизирующего и гепатопротекторного, но и антиатерогенного влияния Таи на показатели липидного обмена. Перечисленные изменения, так же как и продемонстрированное нами увеличение уровня восстановленного глутатиона и KoASH, снижение соотношения в печени АРУЦ/ААК (рис.11), конкурирующих за общие транспортные системы при их поступлении в клетку, указывают на наличие Мембранстабилизирующего, антиокислительного действия Таи. Наличие связи между концентрациями фосфоэтаноламина и Таи было ранее продемонстрировано для плазмы крови и ткани печени, а также отделов головного мозга при стрессе [177].
200-
%% к контролю
100 ■- < ro -в- О о -в- ш О о X со -в- о о е со I -в- о 0> -в- О со -в- О о -в- О сч о Го о си о -в- о < О s I-ro к Рис 12. Антиоксидантная активность, содержание общих фосфолипидов и их фракций, уровни ЭА, ФЭА, KoASH, GSH и соотношение АРУЦ/ААК в печени крыс через 30 минут после в/бр введения таурина (650 мг/кг) %% к контролю Л
100 -- о О Рис 13. Активность ферментов, регламентирующих активность ЦТК (ГДГ, ПДГ, 2-ОГД) и деградацию цистеина (ЦО, ЦТ, ЦД), а также уровень 2-ОГ, ТДФ и соотношения митоходриальных NAD+/NADH в печени крыс через 30 минут после в/бр введения таурина (650 мг/кг) 3.2.2.2 Длительное парентеральное введение Ежедневное внутрибрюшинное введение Таи в дозе 1/10 LD5o вызвало к 1 суткам эксперимента снижение в печени концентрации глутамина, а также абсолютно незаменимой аминокислоты лизина. Кроме этого, повышались уровни аланина и тирозина (рис. 14). Одновременно в печени экспериментальных животных происходило снижение активностей митохондриальной малатдегидрогеназы (MDHm) и цитоплазматической изоцитратдегидрогеназы (ГОНс). Кроме того, наблюдалось повышение уровня лактата (рис.14). На этом фоне в плазме крови опытных животных отмечены менее выраженные сдвиги в содержании исследованых соединений: достоверно увеличились лишь концентрации аланина и пролина (рис.15). К 3 суткам эксперимента в печени животных, получавших Таи, повысилась активность PDH, что указывает на активацию процессов декарбоксилирования пирувата; увеличилась активность цитоплазматической малатдегидрогеназы (MDHc), определяющей NAD-зависимое превращение малата в оксалацетат в цитоплазме гепатоцитов, а также сохранился повышенный уровень лактата (рис. 14). При этом в печени на 1 - 3 сут наблюдалась положительная корреляцонная зависимость между активностью цитоплазматической MDH и уровнем Таи. Примечательно, что при этом значительно снижаются концентрация структурного аналога Таи — J3-Ala (рис. 14) и возникают высокодостоверные (г > 0,90) положительные корреляции между уровнем Таи и его структурного аналога — фосфоэтаноламина в печени, в уровнях Таи и лизина — в плазме крови и отрицательные — между содержанием Таи и активностью ALT в плазме крови. Вышеперечисленное, а также снижение уровня глюкозы на 8 сут, может свидетельствовать об активации процессов гликолиза в печени животных на фоне дополнительного введения Таи. Кроме того, снижение на 1 и 3 сутки соотношения активностей митохондриальной и цитоплазматической форм MDH свидетельствует об ингибировании процессов глюконеогенеза на ранних сроках внутрибрюшинного введения Таи [204]. К 8 и 15 суткам тенденции изменений в уровнях аминокислот и активностях ферментативных реакций, возникшие в более ранние сроки эксперимента, практически сохраняются, кроме этого, на 8 сутки наблюдалось достоверное повышение уровня Таи в печени и плазме крови, а на 15 сутки в печени возникла отрицательная корреляция Таи -Р~А1а. Такая фазовость уровня Таи может объясняться насыщением систем его конъюгации с желчными кислотами, в то время, как в более поздние сроки могут включаться дополнительные адаптивные механизмы поддержания него концентрации. Па этом фоне в плазме крови отчетливо прослеживается увеличение концентрации незаменимых аминокислот Thr и Lys, в печени — снижение уровней глюкозы (GL). Па 15 сутки в плазме крови и печени повышалась концентрация аланина (рис. 14,15), что, вероятно, может свидетельствовать об активации к концу эксперимента процессов глюконеогенеза под действием вводимого Таи. Обращает на себя внимание появление в печени на 15 сутки эксперимента положительной корреляции в уровнях Таи и метионина, что может служить доказательством усиления синтеза эндогенного Таи при его дополнительном введении в течении длительного срока, и появление отрицательной корреляции в уровнях Таи и цистатионина, что, вероятно, свидетельствует о уменьшении относительной значимости других путей превращения метионина. Так уменьшение скорости образования цистатионина может привести в свою очередь и к усилению относительной значимости других путей превращения серина. В пользу последнего предположения говорит появление положительной корреляционной зависимости (г = 0,87) между уровнями Таи и глицина, хотя повышение уровня последнего не было достоверным. На фоне дополнительного внутрибрюшинного введения Таи значительно возросла его концентрация в скелетной мышце (рис.17). Последнее особенно характерно для 3 суток эксперимента, когда в опыте возникают высокодостоверные отрицательные корреляции между уровнями Таи, глицином и (3-А1а. Одновременно наблюдалось обеднение аминокислотного пула сердечной мышцы (снижение концентраций треонина, аланина, метионина, лейцина, лизина) (рис.16) и, в меньшей степени, скелетной мышцы (рис.17), что не наблюдалось в более поздние сроки. К 15 суткам эксперимента в скелетной мускулатуре на фоне высокого содержания Таи снижались концентрации глицина, аспартата и структурного аналога и транспортного антагониста Таи — |3-А1а. Примечательно, что к этому сроку опыта имеется тенденция к снижению содержания Таи в сердечной мышце по сравнению с контролем (рис.16), что подтверждает существование автономных систем транспорта и превращений Таи в сердце. В целом мозге опытных животных дополнительное внутрибрюшинное введение Таи на ранних сроках (1 и 3 суток) от начала эксперимента повысило активность PDH и GDH и ингибировало на 3 сутки ALT и цитоплазматическую MDH (табл. 23). 200 -| □ 1 пооёё D3 пооТё ВвпбоТё П15п66Гё
Рис. 14. Изменения уровней свободных аминокислот и их производных, субстратов гликолиза и активностей ферментов в печени крыс в динамике ежедневного внутрибрюшинного введения Таи в дозе 1/10 LD50
]таурин 1 сут. Iтаурин 8 сут. таурин 3 сут. таурин 15 сут. р < 0,05 по отношению к контролю соответствующего срока Рис.15 Содержание свободных аминокислот, их производных, концентрации субстратов и продуктов гликолиза QlM), а также активность реакций переаминирования (ц. моль/мл/мин), регламентирующих превращения аминокислот в плазме крови крыс в динамике ежедневного внутрибрюшинного введения таурина в дозе 1/10 LD5o (650 мг/кг массы тела)
таурин 1 сут. таурин 8 сут. I таурин 3 сут. I таурин 15 сут. — р < 0,05 по отношению к контролю соответствующего срока Рис. 16 Содержание свободных аминокислот и их производных в сердце крыс в динамике ежедневного внутрибрюшинного введения таурина в дозе 1/10 LD5o (650 мг/кг массы тела ), |Х моль/г
— p < 0,05 по отношению к контролю соответствующего срока Рис.17 Содержание свободных аминокислот и их производных в скелетной мышце крыс в динамике ежедневного внутрибрюшинного введения таурина в дозе 1/10 LD5o (650 мг/кг массы тела), |1 моль/г Таблица 23 Концентрации субстратов и продуктов гликолиза, активность ферментов цикла трикарбоновых кислот а также реакций, регламентирующих превращения аминокислот в мозге крыс в динамике ежедневного внутрибрюшинного введения таурина в дозе 1/10 LD50 (650 мг/кг массы тела ), \i моль/г/мин
* - p < 0,05 по отношению к контролю соответствующей группы 3.2.2.3 Длительное субконъюктивальное введение Концентрация Таи в печени и плазме крови при его субконъюнктивальном способе введения (ежедневно, в течение 15 дней, в суммарной дозе 650 мг/кг массы) не изменилась. Однако уже в первые сутки эксперимента в печени увеличилось содержание LA, в плазме крови — аланина и (З-Ala и снизилось — изолейцина (рис.18,19). Наиболее выраженные изменения при этом способе введения в периферических тканях отмечены к 3 и 8 суткам от начала эксперимента и касаются, в первую очередь, незаменимых и заменимых гликогенных аминокислот. В плазме крови на этих сроках произошло обогащение фонда незаменимых аминокислот (треонин, валин и изолейцин), увеличились концентрации серусодержащих аминокислот (метионин и цистин), АРУЦ (валин, лейцин), а также дикарбоновых аминокислот (глицин, глутамат). Однако, на 15 сутки наблюдалась полная нормализация всего фонда свободных аминокислот в плазме крови (рис.18). Такая фазовость в изменениях уровней аминокислот вероятно можно объяснить изменением процессов их деградации или распада белка. На этом фоне в печени опытных животных возникают высокодостоверные положительные коррелятивные связи между содержанием Таи и уровнем его структурного аналога фосфоэтаноламина (рис.19). В плазме крови опытных животных при субконъюнктивальном способе введения его концентрация высокодостоверно коррелирует с содержанием большинства гликогенных аминокислот. Значительных изменений в концентрациях аминокислот в печени при этом не наблюдалось: на ранних сроках снизилось содержание фенилалнина, а на 15 сутки повысился уровень лейцина и понизился — этаноламина (ЕА) (рис.18). На 15 сутки в печени, кроме того, снизились активности ALT и цитоплазматической ЮН (рис.19).
■ 3 суток ■ 8 суток □ 15 суток Tau Thr Gly Ala Val Cys Met lie Leu —>
Субконъюнктивальное введение Таи незначительно отразилось на структуре фонда свободных аминокислот в скелетной мускулатуре — содержание Таи не изменилось практически на всех сроках, и только на 3 сутки снизилось содержание глутамата, а к 8 суткам повысился уровень глицина (рис.21). При этом возникали высокодостоверные положительные корреляции между концентрациями Таи, глутамин и этаноламин (г>0,90). В сердечной мышце, также, как и в скелетной, субконъюнктивальное введение Таи не изменило его уровень и привело к увеличению на 1 сутки уровня глицина, на 3 сутки — аспартата, треонина, цистина и орнитина (рис.20). На 15 сутки эксперимента в этих структурах произошла полная нормализация всего фонда аминокислот. Таким образом, полученные данные доказывают, что субконъюнктивальное введение Таи на протяжении 15 суток в суммарной дозе 650 мг/кг практически не влияет на процессы формирования аминокислотного фонда в периферических тканях — печени, скелетной и сердечной мышц; наблюдаемые фазные изменения в уровнях свободных аминокислот в плазме крови исчезают к 15 суткам.
Таи urea Asp Thr Ala Leu Туг Phe EA His ALT LA IDHc IDHm Рис. 19. Содержание свободных аминокислот, их производных, концентрации субстратов и продуктов гликолиза ((1моль/г), активность ферментов цикла трикарбоновых кислот и реакций, регламентирующих превращения аминокислот ((хмоль/г/мин) в печени крыс в динамике ежедневного субконъюнктивального введения таурина в дозе 1/10 LD50 (650 мг/кг массы тела) (* - р < 0,05по отношению к контролю соответствующего срока)
таурин 1 сут. таурин 3 сут. таурин 8 сут. таурин 15 сут. Рис.20. Содержание свободных аминокислот и их производных в сердце крыс в динамике субконъюнктивального введения таурина в суммарной дозе 1/10 LD5o (650 мг/кг массы тела ), (Д, моль/г (* - р < 0,05 по отношению к контролю соответствующего срока)
иаурин 1 сут. иаурин 8 сут. иаурин 3 сут. 1таурин 15 сут.
Таи Thr Ser Glu Gly Ala Cys Met bAla Рис.21. Содержание свободных аминокислот и их производных в скелетной мышце в динамике субконъюнктивального введения таурина в суммарной дозе 1/10 LD5o (650 мг/кг массы тела ), ц, моль/г (* - р < 0,05 по отношению к контролю соответствующего срока) Субконъюнктивальное введение Таи приводило, в некоторой степени, к схожим изменениям в целом мозге: на 1 сутки от начала эксперимента активация GDH сопровождалась возникновением положительных корреляций с цитоплазматической ШН, а к 15 суткам на фоне активации митохондриальной ШН, лимитирующей функциональную активность ЦТК (табл. 24), возникали положительные корреляции между активностью ее цитоплазматической формы и процессами переаминирования (активностями AST и ALT). Таблица 24 Концентрации субстратов и продуктов гликолиза, активность ферментов цикла трикарбоновых кислот а также реакций, регламентирующих превращения аминокислот в мозге крыс в динамике субконъюнктивального введения таурина в суммарной дозе 1/10 LD50 (650 мг/кг массы тела ), |1 моль/г/мин
- p < 0,05 по отношению к контролю соответствующей группы 3.3. Нейрэффекторное действие таурина и процессы формирования фонда нейроактивных аминокислот и биогенных аминов в головном мозге 3.3.1 Парентеральное введение таурина Выбор отделов головного мозга с различной метаболической ориентации для выяснения механизмов действия Таи на процессы формирования фонда нейроактивных аминокислот и биогенных аминов в центральной нервной системе обусловлен тем, что гипоталамус является структурой с наиболее высокой проницаемостью гематоэнцефалического барьера (что может означать высокую функциональную активность систем активного транспорта). Кроме этого в гипоталамусе представлены практически все трансмиттерные системы, в том числе значительная часть серотонинергических нейронов [205]. Средний мозг — структура, содержащая основную часть тел дофаминергических нейронов (нигростриального пути), а также значительную часть рецепторов тормозного типа. Выбор стриатума обусловлен низкой активностью МАО в этой структуре, что предполагает наличие в ней постоянно высокой концентраций моноаминов (особенно дофамина) по отношению к продуктам их деградации. Кроме этого в стриатуме наиболее высока концентрация таурина [5, 6, 23]. Исследование влияния однократного внутрибрюшинного введения Таи (1/10 LD5o на 30 мин) на процессы формирования пула нейроактивных аминокислот и биогенных аминов в отделах мозга выявило значительное (более чем в 2 раза) снижение в гипоталамусе опытных животных концентрации J3-Ala — структурного аналога Таи (рис.22), что подтверждает конкурентные взаимоотношения этих соединений при транспорте в мозг. Абсолютные значения концентраций Таи, константы его связывания и скорость транспорта в отделах головного мозга разнятся и не коррелируют между собой [39]. Это предполагает наличие нескольких изоформ Tau-переносящих белков и различный региональный уровень эндогенных биологически активных соединений, конкурирующих за общие с Таи системы транспорта. Концентрация природных аналогов Таи (|3-А1а, GAB А) значительно выше в гипоталамусе по сравнению с другими отделами. Этим, возможно, и объясняется отсутствие повышения уровня Таи в гипоталамусе опытных животных. Стабильный уровень Таи на фоне нагрузки, несмотря на высокую проницаемость сосудистой сети гипоталамуса, свидетельствует, прежде всего, о функциональной значимости Таи в этом отделе ЦНС и существовании активных механизмов поддержания его концентрации. Отделить метаболическую функцию аминокислот от медиаторной невозможно, поскольку нельзя определить из каких пулов (компартментов) произошло их освобождение. Однако, ослабление в опыте положительной коррелятивной связи уровней Glu и Asp с их амидами может расцениватся как нарушение процесса инактивации возбуждающих аминокислот. Характерно, что соотношение суммы тормозных медиаторов (Таи + GABA) и суммы возбуждающих (Glu + Asp) уменьшается (рис.22). Концентрация катехоламинов (NE и DA) при этом уменьшается, что свидетельствует об угнетении процессов гидроксилирования Туг. Положительная корреляция Туг - Таи в гипоталамусе опытных животных становится недостоверной, однако на фоне уменьшения уровня продукта окисления дофамина — DOPAC, возникает прямая коррелятивная связь в содержании DA - Таи (рис.22). Анализ сложившейся в гипоталамусе метаболической ситуации позволяет предположить, что наряду с ингибированием синтеза катехоламинов, происходит адаптивное снижение их катаболизма. Тем самым подтверждается влияние Таи на координацию взаимодействия Glu-, GABA-, и катехоламинергических систем. Дисбаланс пула тормозных медиаторов на этом фоне сопровождается активизацией процессов образования 5-НТ: уровень последнего при введении Таи увеличивается и положительно коррелирует с содержанием Тгр. В целом, однако, в данной экспериментальной ситуации в гипоталамусе возникает относительное обеднение пула тормозных медиаторов, чем по-видимому, могут объяснятся возникающие в эксперименте и на практике случаи амбивалентных эффектов Таи в ЦНС. В стриатуме содержание Таи на фоне нагрузки выросло почти на 30% (рис.22), что прежде всего говорит о большой проницаемости гематоэнцефалического барьера для этого соединения в этих структурах мозга по сравнению с гипоталамусом. При этом увеличение уровня GABA в стриатуме, по-видимому, объясняется связыванием накопившимся Таи цитоплазматического Са2+ на нейрональной мембране и активацией в результате этого глутаматдекарбоксилазы. В стриатуме опытных животных, кроме этого, выросло содержание DA (рис.22), что с возникновением отрицательных корреляций Таи с NE и метаболитами DA — DOPAC и HVA может свидетельствовать об угнетении процессов инактивации DA. Функционально эти сдвиги могут служить основой тормозного действия Таи и стабилизации им моторной функции ЦНС. Оценка создавшейся в исследованных отделах мозга ситуации позволяет предположить существование различной степени деформации метаболических процессов, обусловленной, прежде всего, индивидуальными для структур мозга особенностями транспорта экзогенного Таи кровью. Для сформировавшегося мозга активный транспорт Таи против градиента концентрации является одним из наиболее значимых в поддержании стабильного уровня этого соединения.
Поиск по сайту: |


 Рис.11 Изменения уровней свободных аминокислот и их производных в печени и плазме крови крыс, соотношения митоходриальных НАД+/НАДН, активностей PDH и GDH, содержания 2-OG в печени через 30 мин после однократного в/бр введения Таи (650 мг/кг)
Рис.11 Изменения уровней свободных аминокислот и их производных в печени и плазме крови крыс, соотношения митоходриальных НАД+/НАДН, активностей PDH и GDH, содержания 2-OG в печени через 30 мин после однократного в/бр введения Таи (650 мг/кг)
















 Tau Glu Gin Ala Tyr I —Ala |i Lys PDH MDHc MDHm LA GL
Tau Glu Gin Ala Tyr I —Ala |i Lys PDH MDHc MDHm LA GL 200 180 160 140
200 180 160 140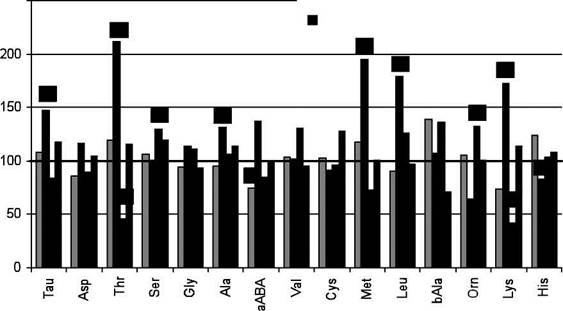 25U
25U таурин 1 сут. таурин 3 сут. таурин 8 сут. таурин 15 сут.
таурин 1 сут. таурин 3 сут. таурин 8 сут. таурин 15 сут. -г% К КОНТРОЛЮ
-г% К КОНТРОЛЮ □ 1 сутки
□ 1 сутки Рис.18 Изменения уровней свободных аминокислот и их производных в плазме крови крыс в динамике ежедневного субконъюнктивального введения таурина в суммарной дозе 1/10 LD5o
Рис.18 Изменения уровней свободных аминокислот и их производных в плазме крови крыс в динамике ежедневного субконъюнктивального введения таурина в суммарной дозе 1/10 LD5o
 16С 140
16С 140 300-
300-