|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Розлади програми управління
Бенек (Benecke, 1989) розрізняє рухові програми і плани рухів. Згідно цієї термінології, рухові програми — це стереотипні черговості команд в ЦНС, які визначають порядок іннервації різних м'язів. Плани рухів координують декілька рухових програм і пристосовують їх до необхідних умов. Рухова програма простого руху руки вперед передбачає твердо встановлену черговість в активації агонистов і антагоністів, яку можна вивести на ЕМГ (електроміограму) і отримати характерний трифазний патерн (Benecke et al., 1985). Така послідовність м'язових активацій приводить до плавного руху, що характеризується єдиним максимумом швидкості. Змінюючи тимчасові інтервали між активацією агонистов і антагоністів і модулюючи силу окремих м'язових активацій, можна досягти в рамках однієї і тієї ж рухової програми рухів різної швидкості і амплітуди (Berardelli et al., 1996, Plamondon, 1995a, b). Це пристосування рухової програми до різних умов називається параметризацією. Розлад параметризації рухових програм має місце, наприклад, при синдромі Паркінсона. Синдром Паркінсона — це розлад довільних і мимовільних рухів з уповільненням спонтанних і довільних рухів (брадикинезия), супроводжуване ригідністю і тремором у спокої (Scholz & Oertel, 1993). Для пацієнтів з синдромом Паркінсона характерні маскоподібний вираз обличчя, монотонна манера говорити і човгаюча хода маленькими кроками. Аналіз ЕМГ показує, що у пацієнтів з синдромом Паркінсона навіть прості рухи рук відрізняються трифазним патерном. Але при здійсненні руху до предметів, що знаходяться на різному віддаленні, виявляється, що окремі компоненти ЕМГ не адекватні необхідній дальності руху, внаслідок чого виходять повільні і дуже короткі рухи (Berardelli et al., 1986). Таким чином, рухова програма ще зберігається, проте вона не може забезпечити адекватності різних рухів. Разом з цим у пацієнтів з синдромом Паркінсона мають місце і розлади координації різних рухових програм (див. прим. 25.2.1).
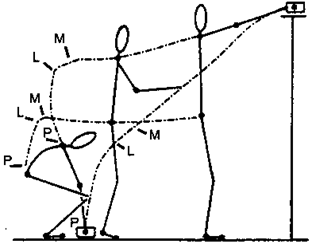
Примітка 25.2.1. Порушена координація рухових програм у пацієнтів з синдромом Паркінсона Коли ми беремо предмет з підлоги і ставимо на полицю, що знаходиться на віддалі, ми повинні перейти із зігнутої пози у випрямлену, зробити декілька кроків і підняти руку, щоб покласти цей предмет на полку. У цьому русі поєднуються випрямний рух (P), горизонтальний рух (L), що продовжується, і підняття руки (M) (мал. 25.2.1).
Мал. 25.2.1. Ілюстрація рухового завдання. Пояснення в тексті (висловлюється по Ingvarsson et al., 1986)
Щоб проаналізувати координацію цього складного руху, Інгварссон і ін. (Ingvarsson et al., 1986) зняли руху спеціальною відеосистемою і потім розрахували траєкторії і швидкість для предмету і суглобів, що беруть участь. Мал. 25.2.2 показує кут підйому руки (ALFA) і горизонтальну (Vx) і вертикальну (Vy) швидкість предмету в ході руху.
Мал. 25.2.2. Порівняння руху, що відбувається: А — здоровий випробовуваний, і В — пацієнт з синдромом Паркінсона. Пояснення в тексті (висловлюється по Ingvarsson et al., 1986)
Мал. 25.2.2А показує рух здорового випробовуваного. Спочатку кут підйому і горизонтальна швидкість об'єкту постійні; об'єкт рухається тільки у вертикальному напрямі. У цій фазі має місце тільки частковий випрямний рух від стегна (P). Вже через декілька мілісекунд починається рух ходьби (L). Початок цього руху позначений «L». Горизонтальний і вертикальний рухи протікають приблизно паралельно. Ще до того, як горизонтальний рух (ходьба) досягає максимуму швидкості, починається рух підняття руки (M). Ці три компоненти руху можна бачити і на мал. 25.2.1. Пунктирні лінії відтворюють траєкторію плеча, стегна і об'єкту. У здорових випробовуваних три рухи стартують один за одним з коротким латентним періодом і на широкому відрізку сукупного руху виконуються паралельно. Результатом цій досить добре настроєній координації трьох рухів є те, що об'єкт плавно і по практично лінійній траєкторії транспортується від початкової точки до місця мети. Абсолютно інакше виглядає рух у пацієнтів з синдромом Паркінсона (мал. 25.2.2В). Тут рух (ходьба) (Vx) починається тільки після того, як випрямний рух (Vy) вже майже завершився. Рух підняття руки (M), у свою чергу, починається тільки після закінчення руху ходьби. Три компоненти руху у пацієнтів з синдромом Паркінсона майже повністю розузгодили. Результат — виразне уповільнення руху. Таким чином, істотний аспект рухового розладу у пацієнтів з синдромом Паркінсона слід бачити в тому, що при виконанні складних рухів порушена координація окремих рухових програм. ---
Розлади регуляції
В більшості випадків рухи, адекватні навколишньому світу, здійснюються завдяки зору. Ця зрительно-моторная координація представляється нам в нормі чимось само собою зрозумілим і тривіальним. Проте її розвиток залежить від наявності певних умов (ср. Heuer, 1983, S. 26-29) і може бути порушене. Природність деяких функцій, що здається, таких як зрительно-моторная координація, очевидно означає не те, що система, що реалізовує ці функції, проста і здорова, а лише те, що її активність не супроводжується свідомим переживанням. Дуже специфічне зрительно-моторное розузгодження має місце при оптичній атаксії, яка зазвичай є наслідком пошкодження задньої області тім'яних доль (Newcombe & Ratcliff, 1989). Пацієнти не в змозі схопити рукою який-небудь об'єкт. Залежно від локалізації мозкового пошкодження цей розлад може торкнутися тільки однієї руки, тільки одну половину особи або тільки одну специфічну комбінацію руки і половини особи (Rondot et al., 1977). Оптична атаксія не є моторним розладом у вузькому сенсі слова. В протилежність кінетичній атаксії рухи самі по собі плавні і не порушені (Jakobson et al., 1991). Немає вона і порушенням правильного сприйняття, оскільки пацієнти більш менш в змозі описати форму, місцеположення і ідентичність предмету (Jeannerod, 1988). Візуальна дія і моторна компетентність сохранны, проте візуальна інформація, очевидно, не може використовуватися для моторного управління. Цікаво, що буває і зворотний феномен: Гудейл і Мілнер (Goodale & Milner, 1992) повідомляють про пацієнтку з темпорально-окципитальным мозковим пошкодженням, яка не розпізнавала величину і форму об'єкту, але, беручи цей об'єкт, розкривала руку рівно настільки, скільки потрібне відповідно до його форми і величини. Як і Унгерлейдер і Мішкин (Ungerleider & Mishkin, 1982), які розрізняють в мозку дві окремі візуальні системи, Гудейл і Мілнер (Goodale & Milner, 1992) пропонують підрозділяти візуальну систему на систему сприйняття і систему дії. Темпорально-окципітальная система служить для ідентифікації об'єктів, парієнтальна — надає візуальну інформацію для управління рухом. Відповідно у разі оптичної атаксії є селективний розлад парієнтальної системи.
Розлади координації
У реалізації певного руху найчастіше бере участь ряд м'язів, які повинні скорочуватися в певній послідовності і з певною інтенсивністю. При цьому можна говорити про деяку толерантність до помилок, оскільки частіше всього один і той же результат руху може досягатися за рахунок різних патернів скорочення, але, природно, є і межі, за межами яких рух не вдається. Як відбувається оволодіння трансформацією тіла і що для цього потрібний, можна пояснити на прикладі листи. Бажаний результат руху залежить від певної форми руху, наприклад від зображення певного штриха. При цьому доводиться мати справу тільки з двома просторовими вимірюваннями (якщо нехтувати рухом олівця, коли, починаючи писати, його опускають на папір). Але що стосується задіяних суглобів, то тут число вже більше. Їх дрібні і швидкі рухи навряд чи доступні самоспостереженню. Координація працюючих м'язів зрештою здійснюється без якої-небудь участі нашої свідомості. Одне можна затверджувати напевно: подібна система з багатьма компонентами, від яких вимагається досить точна спільна робота, легко схильна до порушень. У дітей або коли пишуть не домінантною рукою, втім, спостерігається спрощення трансформації тіла і тим самим спрощення координації: дистальні пальцеві суглоби або зап'ястя ціпеніють і лист здійснюється швидшим за рахунок проксимальных суглобів. Однією з простих форм розладу координації може бути свого роду «надмірна активність», коли замість тонко узгодженої поєднаної роботи різних м'язів зростає м'язова напруга в цілому, і на цьому високому рівні його можна модулювати лише насилу. Типова картина при цьому — одночасне скорочення м'язів-антагоністів. Це явище має місце при писальному спазмі. Писальний спазм, або графоспазм, — це руховий розлад, при якому рухи при письмі характеризуються надмірною напругою м'язів пальців, кисті і передпліччя або всієї руки. Багато пацієнтів починають писати, ставлячи кисть і руку в незвичні позиції, прикладають всі сили, щоб тримати олівець, і сильно спираються на стіл. Під час листа м'язова напруга найчастіше ще більше зростає, і деякі пацієнти не можуть написати більше декількох слів. Дивно, що при цьому рідкісному розладі трудності зазвичай обмежуються листом. Віндгассен і Лудольф (Windgassen & Ludolph, 1991) повідомляють про одного хірурга з важким писальним спазмом, який ніколи не міг написати більше однієї короткої пропозиції, але без зусиль робив складні хірургічні операції. Чим обумовлюється цей високоспецифічний розлад — питання спірне. В даний час більшість дослідників припускають, що мова йде про якомусь органічному розладі, патофізіологічні основи якого поки не відомі (Sheehy & Marsden, 1982; Ludolph & Windgassen, 1992). Детальний аналіз рухів у пацієнтів з писальним спазмом проте показує, що разом з явно порушеними рухами у всіх пацієнтів деякі рухи зберігаються (Mai et al., 1996). Значне поліпшення здатності писати досягається до того ж за допомогою відповідного тренінгу (Mai et al., 1994). Разом узяті, ці дані наводять на думку, що при писальному спазмі мова йде про функціональному, а не про органічний руховий розлад (ср. розділ 25.1). Ще один приклад розладу координації — це кінетична атаксія (розлад цілеспрямованих рухів). Пацієнти з атаксією не досягають мети руху, ініціація рухів сповільнена, хід руху порушений: замість плавного руху робиться багато коротких приватних рухів (ср. мал. 25.1.1, розділ 25.1). Характерним для кінетичної атаксії є так званий интенционный тремор. Це низькочастотний, нерегулярний тремор, який наступає в кінці цілеспрямованого руху. Огляд різних розладів при атаксії дають Дінер і Діхганс (Diener & Dichgans, 1992). Атактичні розлади найчастіше наступають у зв'язку з якимсь пошкодженням мозочка, і інтерпретація атактичного розладу сильно залежить від розвитку теорій про функцію мозочка. У історії дослідження мозочка дискусія розгорілася навколо питання: за що ответствен мозочок — за модуляцію простих рухів або ж за координацію складних? Флоуренс (Flourens, 1824), грунтуючись на своїх дослідах з голубами, прийшов до висновку, що мозочок координує рухи. Лючиані (Luciani, 1915) відкинув теорію Флоуренса, після того, як йому довелося спостерігати собаку, який, не дивлячись на те що у неї була видалена половина мозочка, виконувала чудово скоординовані плавальні рухи. В світлі поглядів Лючиані сформувалася думка, що мозочок надає на моторику виключно модулюючу дію. Істотною функцією мозочка почали вважати гальмування активності усередині рефлекторних дуг. Було висунуто припущення, що розлад цього гальмування несе відповідальність за атактичний тремор, який розглядається як ефект розгойдуючої осциляції в рефлекторних дугах (Glaser & Higgins, 1966). Проте виникає питання: чому такий складний орган, як мозочок, що містить більше нейронів, чим решта всього мозку, необхідний для виконання такої функції, яка в електричних ланцюгах виконується простими структурами опору? Деякі учені (наприклад, Thach, Goodkin & Keating, 1992) пропонують модель функціонування мозочка, яка віддає належне складності цього органу і в якій враховуються нові дані анатомії і фізіології мозочка. Згідно цієї моделі, мозочок регулює специфічна для завдання поєднана дія різних м'язів, що беруть участь в русі (так звані синергии), і здійснює специфічну ж для завдання модуляцію рефлексів. Пошкодження мозочка приводить до часткової втрати цієї синергии і порушує її повторне придбання. Комбінацією цих двох функцій мозочка можна пояснити (при їх пошкодженні) порушення координації і тремор, спостережувані при атаксії. Проте і в цьому випадку прийнято вважати, що клінічна картина зазвичай стає абсолютно зрозумілою тільки при додатковому обліку наслідків вторинних адаптаційних процесів. Інакше навряд чи можна було б пояснити прогредиентное перебіг деяких посттравматичних атаксій, а також той факт, що у деяких пацієнтів вираженість атаксії варіює залежно від «небезпеки» ситуації (ср. мал. 25.1.1, глава 25.1; Mai et al., 1995).
Література
Baxter, P. S. (1995). Attention-deficit hyperactivity disorder in children. Current Opinion in Pediatry, 7, 381-386. Benecke, R. (1989). The pathophysiology of Parkinson's Disease. In N. P. Quinn & P. G. Jenner (Hrsg.), Disorders of Movement. Clinical Pharmacological and physiological aspects. London: Academic Press, 59-72. Benecke, R., Meinck, H. M. & Conrad, B. (1985). Rapid goal-directed elbow flexion movements: limitations of the speed control system due to neural constraints. Experimental Brain Research, 59, 470-477. Berardelli, A., Dick, J. P., Rothwell, J. C, Day, B. L. & Marsden, C. D. (1986). Scaling of the size of first agonist EMG burst during rapid wrist movements in patients with Parkinson's disease. Journal of Neurology, Neurosurgery and Psychiatry, 49, 1273-1279. Berardelli, A. Hallett, M., Rothwell, J. C., Agostino, R., Manfredi, M., Thompson, P. D. & Marsden, C. D. (1996). Single-joint rapid arm movements in normal subjects and in patients with motor disorders. Brain, 119,661-674. Bliss, J. (1980). Sensory experiences in Gilles de la Tourette syndrome. Archives of General Psychiatry, 37, 1343-1347. Brown, C. S. (1991). Treatment of attention deficit hyperactivity disorder: а critical review. Developments in clinical pharmacology, 25, 1207-13. Caine, E. D., McBride, M. C., Chiverton, P., Bamford, K. A., Rediess, S. & Shiao, J. (1988). Tourette's syndrom in Monroe county school children. Neurology, 38, 472-475. Churchland, P. M. (1984). Matter and conscious experience. Cambridge, Mass.: MIT Press. Conners, C. K. & Wells, K. C. (1986). Hyperkinetic children. A neuropsychosocial approach. London: Sage. Crisp, A. H. & Moldofsky, H. (1965). A psychosomatic study of writer's cramp. British Journal of Psychiatry, 111, 841-858. Cruse, H., Dean, J., Heuer, H., & Schmidt, R. A. (1990). Utilization of sensory information for motor control. In O. Neumann & W. Prinz (Eds.), Relationships between perception and action. Current approaches (pp. 43-79). Berlin: Springer. Della Sala, S., Marchett, C. & Spinnler, H. (1994). The anarchic hand: а fronto-mesial sign. In F. Boller & J. Grafman (Hrsg.), Handbook of Neuropsychology (Vol. 9, pp. 33-255). Amsterdam: Elsevier. Diener, H. C. & Dichgans, J. (1992). Pathophysiology of cerebellar ataxia. Movement Disorders, 7, 95-109. Flourens, P. (1824). Recherches experimentales sur les propriйtйs et les fonctions du systeme nerveux, dans les animaux vertйbres. Paris: Crevot. Glaser, G. H., Higgins, D. C. (1966). Motor stability, stretch responses, and the cerebellum. In R. Granit (Eds.) Muscular Afferents and Motor Control. Proc. Nobel Symp., (Vol. 1, pp. 121-138). Stockholm: Almquist & Wiksell. Goldberg, G. & Bloom, K. (1990). The alien hand sign: localization, lateralization and recovery. American Journal of Physical Medical Rehabilitation, 69, 228-238. Goldenberg, G. (1995). Imitating gestures and manipulating а mannikin — the representation of the human body in ideomotor apraxia. Neuropsychologia, 33, 63-72. Goldenberg, G., Wimmer, A., Holzner, F. & Wessely, P. (1985). Apraxia of the left limbs in а case of callosal disconnection: the contribution of medial frontal lobe damage. Cortex, 21, 135-148. Goodale, M. A. & Milner, A. D. Separate visual pathways for perception and action. Trends in Neuroscience, 15, 20-25. Hechtman, L. (1994). Genetic and neurobiological aspects of attention deficit hyperactive disorder: а review. Journal of Psychiatry and Neuroscience, 19, 193-201. Heilman, K. M., Voeller, K. K. & Nadeau, S. E. (1991). A possible pathophysiologic substrate of attention deficit hyperactivity disorder. Journal of Child Neurology, 6, 76-81. Hermsdцrfer, J., Mai, N., Spatt, J., Marquardt, C., Veitkamp, R. & Goldenberg, G. (1996). Kinematic analysis of movement imitation in apraxia. Brain, 119, 1575-1586. Heuer, H. (1983) Bewegungslernen. Stuttgart: Kohlhammer. Heuer, H. (1992). Psychomotorik. In H. Spada (Hrsg.), Lehrbuch Allgemeine Psychologie (2. Auflage, S. 495-559). Bern: Huber. Heuer, H. (1996) Doppeltдtigkeiten. In O. Neumann & A. F. Sanders (Hrsg.), Enzyklopдdie der Psychologie C II 2: Aufmerksamkeit (S. 163-218). Gцttingen: Hogrefe. Heuer, H. & Keele, S. W. (Hrsg.) (1994), Enzyklopдdie der Psychologie C II 3: Psychomotorik. Gцttingen: Hogrefe. Heuer, H. & Prinz, W. (1987). Initiierung und Steuerung von Handlungen und Bewegungen. In M. Amelang (Hrsg.), Bericht ьber den 35. KongreЯ der Deutschen Gesellschaft fьr Psychologie in Heidelberg 1986 (S. 289-295). Gцttingen: Hogrefe. Ingvarsson, P., Johnels, B., Lund, S. & Steg, G. (1986). Coordination of manual, postural, and locomotor movements during simple goal-directed motor tasks in parkinsonian off and on states. Advances in Neurology, 45, 375-382. Jakobson, L. S., Archibald, Y. M., Carey, D. P. & Goodale, M. A. (1991). A kinematic analysis of reaching and grasping movements in а patient recovering from optic ataxia. Neuropsychologia, 29, 803-809. Jeannerod, M. (1988). The Neural and Behavioural Organization of Goal-Directed Movements. Oxford: Oxford University Press. Leckman, J. F., Walker, D. E. & Cohen, D. J. (1993). Premonitory urges in Tourette's syndrome. American Journal of Psychiatry, 150, 98-102. Lee, A, J. (1989). Tics. In N. P. Quinn & P. G. Jenner (Eds.), Disorders of Movement. Clinical Pharmacological and physiological aspects (pp. 495-504). London: Academic Press. Lhermitte, F. (1986). Human autonomy and the frontal lobes. Part II: Patient behavior in complex and social situations: the «environmental dependency syndrome». Annals of Neurology, 19, 335-343. Luciani, L. (1915). The hindbrain. In F. A. Welby (Ьbersetzer): Human Physiology. 467. London: Macmillan. Ludolph, A. C. & Windgassen, K. (1992). Klinische Untersuchungen zum Schreibkrampf bei 30 Patienten. Nervenarzt, 63, 229-243. Mai, N., Marquardt, C. (1994). Treatment of writer's cramp. In C. Faure, P. Keuss, G. Lorette & A. Vinter (Eds.), Advances in handwriting and drawing. A multidisciplinary approach. Paris: Telecom. Mai, N., Marquardt, C. (1995). Analyse und Therapie motorischer Schreibstцrungen. Psychologische Beitrдge, 37, 538-582. Mai, N., Przywara, S., Hermsdцrfer, J. & Marquardt, C. (1995). Behandlung der posttraumatischen Ataxie. In Kuratorium ZNS (Hrsg.) Forschung und Praxis der Neurologischen Rehabilitation — 10 Jahre Kuratorium ZNS. Hamburg: In-Transfer (S. 241-256). Newcombe, F. & Ratcliff, G. (1989). Disorders of visuo-spatial analysis. In F. Boiler & J. Grafman (Eds.), Handbook of Neuropsychology (Vol. 9, pp. 333-356). Amsterdam: Elsevier. Neumann, O. (1987). Beyond capacity: A functional view of attention. In H. Heuer & A. F. Sanders (Eds.), Perspectives on perception and action (pp. 361-394). Hillsdale, N.J.: Erlbaum. Obeso, J. A., Rothwell, J. C. & Marsden, C. D. (1981). Simple tics in Gilles de la Tourette's syndrome are not prefaced by а normal premovement EEG potential. Journal of Neurology, Neurosurgery, and Psychiatry, 44, 735-738. Plamondon, R. (1995a) A kinematic theory of rapid human movements: 1. Movement representation and generation. Biological Cybernetics, 72, 295-307. Plamondon, R. (1995b) A kinematic theory of rapid human movements: 2. Movement time and control. Biological Cybernetics, 72, 309-320. Prinz, W. (1987). Ideo-motor action. In H. Heuer & A. F. Sanders (Eds.), Perspectives on perception and action (pp. 47-76). Hillsdale, N.J.: Erlbaum. Prinz, W. (1992). Why don't we perceive our brain states? European Journal of Cognitive Psychology, 4, 1-20. Rondot, P., de Recondo, J. & Ribadeau Dumas, J. L. (1977). Visuo-motor ataxia. Brain, 100, 355-376. Sanes, J. N., Mauritz, K.-H., Dalakas; M. C. & Evarts; E. V. (1985). Motor control in humans with large-fibre sensory neuropathy. Human Neurobiology, 4, 101-114. Scholz, E. & Oertel, W. H. (1993). Parkinson-Syndrome. In T. Brandt, J. Dichgans & H. C. Diener (Hrsg.), Therapie und Verlauf neurologischer Erkrankungen (2. Aufl., S. 927-968). Stuttgart: Kohlhammer. Sheehy, M. P. & Marsden, C. D. (1982). Writer's cramp - а focal dystonia. Brain, 105, 461-480. Sьss-Burghart, H. (1996). Verhaltenstherapie mit einem 11 Jahre alten Jungen mit der Diagnose Gilles-de-la-Tourette-Syndrom - ein Fallbericht. Verhaltenstherapie, 6, 100-106. Thach, W. T., Goodkin, H. P. & Keating, J. G. (1992). The cerebellum and the adaptive coordination of movement. Annual Review of Neuroscience, 15, 403-442. Watson, R. T. & Heilman, K. M. (1983). Callosal Apraxia. Brain, 106, 391-403. Ungerleider, L. G. and Mishkin, M. (1982). Analysis of Visual Behavior. In D. J. Ingle, M. A. Goodale & R. J. Mansfield (Eds.), Analysis of Visual behaviour (pp. 549-586). Cambridge, M. A.: MIT Press. Windgassen, K. & Ludolph, A. (1991). Psychiatric aspects of writer's cramp. European Archives of Psychiatry and Clinical Neuroscience, 241, 170-176.
Поиск по сайту: |