|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Методы анализа таксономических данных
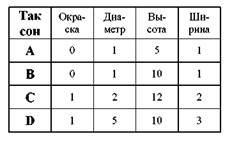
Таксономическое исследование давно вышло за пределы произвольного комбинирования таксонов согласно вкусам и представлениям исследователя. На протяжении последних десятилетий создана мощная методологическая база, которая позволяет в значительной мере объективизировать и стандартизировать работу таксономиста. Современные исследования в области систематики практически не обходятся без применения тех или иных специализированных методов. Типологический метод Типологический метод основан на логическом анализе признаков, присущих организмам. Это – классический метод анализа фенотипических признаков, сложившийся еще в XVIII веке. Построение классификации с помощью типологического метода включает следующие этапы: 1) Все индивидуумы распределяются на группы в соответствии с тем, каким известным типовым (эталонным) образцам они идентичны. Разумеется, в большинстве случаев используются не сами образцы, а их описания в литературе. 2) Производится учет присущих созданным группам признаков и модусов. 3) Проводится оценка значимости всех учтенных признаков с позиций клас- сификации, т.е. взвешивание признаков. Эта процедура является в типологическом методе чисто субъективной и опирается на представления исследователя о ходе эволюции изучаемой группы. В результате всем признакам присваивается определѐнный таксономический вес. 4) В зависимости от присвоенного таксономического веса, признаки разби- ваются на группы, и каждой группе приписывается определенный таксономический ранг. Например, для водорослей постулируется, что пигментный состав хлоропластов характеризует отделы, морфология таллома – классы, структура жгутикового аппарата – порядки, а тип клеточных покровов – семейства. Таким образом, анализируя признаки образца, принадлежащего к определенному виду, исследователь признает, что далеко не все они характеризуют исследуемый таксон именно как вид. Каждый образец несѐт в себе также признаки рода, семейства, порядка или, напротив, разновидности и формы. Сравнивая один вид с другим, ученый сравнивает лишь признаки «видового уровня», относя этот вид к роду – «родовые» признаки, относя к семейству – признаки семейства. Такие комплексы признаков, отличающие друг от друга таксоны данного иерархического уровня называют типами групп. 5) Последний этап типологического анализа – разграничение таксонов. В ходе этого процесса, поочерѐдно, в соответствии с таксономическим весом, учитываются состояния всех признаков типа каждой группы. Вначале анализируются наименее значимые признаки, и организмы распределяются по мелким таксонам. Затем рассматриваются более значимые признаки, и организмы объединяются в более крупные таксоны. В итоге образуется стройная классификация изучаемой группы. Достоинства Типологический метод имеет большую историю и глубокие традиции. Он внѐс в систематику много конструктивных идей, таких как разграничение признаков и их состояний, понятие типа группы, и, конечно же, представление о типовых образцах, на котором строится вся современная биологическая номенклатура. Классификации, созданные типологическими методами, ясны, логичны и необыкновенно удобны в работе, поскольку зачастую построены по принципу определительного ключа. При классифицировании малоизученных, микроскопических организмов типологический метод зачастую оказывается наиболее эффективным. Недостатки Главным недостатком типологического метода является принципиальная субъективность принятого в нем взвешивания признаков. Установить чѐт-кие, обоснованные и универсальные принципы такого взвешивания просто невозможно: как, например, объединить признаки семейства кошачьих и семейства сыроежковых в единую категорию признаков семейства? Итак, с теоретических позиций типологический метод оказывается чрезвычайно слаб, являясь, скорее, не научным методом, а системой строго регламентированных правил, полезных в рутинной работе таксономиста. Эволюционно-систематический метод С начала 1950-х годов таксономическая теория подверглась существенному влиянию школы, получившей в начале название «новая систематика» а затем – «эволюционная систематика». Создатели этой школы, орнитолог Э.Майр и палеонтолог Дж.Г.Симпсон, изучая разнообразие позвоночных животных, пришли к выводу, что описать это разнообразие с помощью стандартных методов и унифицированных принципов абсолютно невозможно. В противоположность типологам, они утверждали, что признак не может обладать универсальной значимостью: в классификации одной группы он важен, а в другой его приходится игнорировать. К таким признакам можно отнести и родство между крупными таксонами. Э.Майр (Mayr, 1953) разработал популяционный аспект нового метода. Согласно его точке зрения, таксономическое исследование должно строится не на изучении типовых образцов, а на сравнении популяций и оценке внутривидовой изменчивости. Дж.Г.Симпсон (Simpson, 1961) изложил палеонтологический аспект учения эволюционной систематики. Он полагал, что выявление филогенеза определенных групп – это полезная, но не основная цель биологической систематики. Главная же ее цель – понять, как взаимосвязаны и сгруппированы организмы с биологической точки зрения. По мнению Дж.Г.Симпсона, классификации должны содержать максимальное количество информации. Классификация и филогения, по мнению Дж.Г.Симпсона, это разные, хотя и взаимосвязанные вещи. Классическая эволюционная систематика стремится внести максимальное количество эволюционной информации в биологические классификации. Формальная классификация является попыткой отразить одновременно генеалогию и степень различий (дивергенции) между родственными организмами. При этом полное соответствие между классификацией и филогенией оказывается невозможным. С использованием этих представлений была разработана своеобразная система анализа таксономических данных, которая реализуется в три основных этапа. 1) Проводится всестороннее исследование группы организмов по всем возможным фенотипическим параметрам: морфофизиологическим, экологическим, биолого-этологическим. По возможности он включает исследование реальных популяций, их структуры, границ, степени изоляции и т. п. Исследуется также филогенез группы, преимущественно по данным палеонтологии. 2) Из накопленного комплекса данных исследователь в каждом конкретном случае выбирает те, которые, по его мнению, обладают наибольшей значимостью. Сходство между организмами и предполагаемое родство между ними рассматриваются при этом как равноценные параметры, и исследователь волен решать, в какой мере генеалогия организмов повлияет на структуру создаваемой классификации. Итак, здесь производится уже знакомое нам взвешивание признаков, в котором, однако, задействован новый параметр – филогенез. 3) На основе выбранных критериев организмы объединяются в таксоны. Если в пределах крупного таксона (например, класса А) выделяется группа (отряд b), приобретшая в ходе эволюции ряд существенных отличий от остальных отрядов данного класса, таксономист вправе выделить данную группу в самостоятельный класс В. При этом, класс А будет считаться ныне существующим предком класса В. Подобное соотношение таксонов, известное как парафилия (см. ниже), признается законным только сторонниками эволюционно-систематического метода. Достоинства Фундаментальные исследования, проведенные сторонниками эволюционно-систематического метода, внесли значительный вклад в изучение высших организмов. Они показали ограниченность формальных типологических классификаций, и взяли на себя титаническую работу всестороннего исследования каждого вида. В изучении малых таксонов высших организмов эво-люционно-систематический метод до сих пор остается приоритетным направлением. Недостатки Колоссальная трудоемкость эволюционно-систематического метода делает его реально применимым лишь в систематике малых таксонов. В работе с надвидовыми группами он теряет «почву под ногам», т. к. лишается важнейших критериев – репродуктивных и популяционно-экологических. В настоящее время не существует методов, которые позволили бы точно оценить признаки групп-«потомков» и выяснить, какие из них заслуживают вынесения за пределы группы-«предка», а какие должны рассматриваться в ее пределах. Поэтому признание парафилетических таксонов влечет за собой значительную субъективизацию таксономического исследования. Эволюционно-систематический метод совершенно не применим в тех случаях, когда непосредственная работа с популяциями и проведение скрещивания невозможно: при работе с микроорганизмами на природном материале, в исследованиях гербариев и коллекций или в тех случаях, когда объект исследования не способен к половому размножению. Следовательно, при- менение этого метода в микологии, альгологии, протозоологии и бактериологии весьма ограничено. Фенетический метод Все фенотипические методы требуют анализа огромного числа признаков. Типологический подход и метод эволюционной таксономии предлагают вычленить среди признаков наиболее значимые. Однако это не единственный способ справиться с обилием информации. Альтернативный путь предлагает нумерическая таксономия. В конце XVIII века французский ботаник М.Адансон (Adanson) заявил, что все признаки организмов являются таксономически равными и должны анализироваться в полном объеме, с привлечением математических методов. Однако длительное время идея М.Адансона считалась неосуществимой, т.к. требовала обработки колоссального количества данных. Лишь в начале 1960-х гг. американский зоолог Р.Сокэл (Sokal) предложил использовать для таксономического анализа средства ЭВМ. Применение компьютера позволило одновременно обрабатывать данные о сотнях видов, оперируя при этом тысячами признаков. Разработанный на этой основе метод первоначально получил название ну-мерическая таксономия. В 1965 г. Э.Майр назвал это направление фене-тикой, что, по-видимому, предпочтительнее, т.к. количественные (нумери-ческие) подходы существуют и в других направлениях систематики. Кроме того, этот метод известен как таксономия Адансона. По мере развития средств математической статистики, были созданы специальные компьютерные программы, которые обеспечили возможность широкого применения фенетического метода. Среди них наибольшее распространение получили Excel, Statistica и PAUP. Благодаря этим программам сложнейшее нумерическое исследование стало в настоящее время доступным практически каждому таксономисту. Построение классификации с помощью фенетического метода включает следующие этапы: 1) На первом этапе производится детальное описание максимально возможного числа количественных (длина, высота, диаметр) и качественных (цвет, форма, орнаментация) характеристик организма. При этом желательно делать множество учетов каждого параметра на основании одного или нескольких образцов данного вида. Для количественных признаков учитываются непосредственные результаты измерений, а для качественных производится их зашифровывание в виде условного цифрового кода. В простейшем случае используется кодировка «0 / 1» т.е. «нет / есть». Если же качественный признак имеет разные проявления, например если окраска может быть серой, черной или коричневой, то вид описывается характеристикой «серый = 1, коричневый = 2, черный = 3». Все полученные таким образом числовые данные заносятся в единую таблицу – первичную матрицу (рис.1). Ее составление производится без участия вычислительных способностей компьютера и представляет собой наиболее трудоемкий этап исследования. 2) Собранные данные, уже с помощью программы, подвергаются статисти- ческой обработке: выясняется, какому типу математического распределения подчиняется разброс проявлений каждого признака, рассчитывается среднее значение изучаемых величин, стандартное отклонение и другие статистические параметры. На этом этапе полезно произвести корреляционный анализ признаков, позволяющий выяснить, насколько сходны их проявления в различных таксонах. Это позволяет оценить своеобразие или наоборот, тождественность поведения данного признака по отношению к другим. Например, признаки 1 и 2 можно считать коррелирующими относительно таксонов А и В если в таксоне А оба признака находятся в состоянии «0», а в таксоне В – в состоянии «1». 3) Все данные первичной матрицы подвергаются процедуре стандартиза- ции (нормализации) – перекодированию всех значений в соответствии с единой шкалой: от -1 до +1. Это позволяет сделать все числа матрицы сравнимыми между собой. Стандартизация проводится по формуле:
где Xn – стандартизованное значение, Х – исходное значение, X – среднее значение параметра Х и D – дисперсия параметра Х. В результате создается вторичная матрица (рис.1) Рис. 1. Матрицы Первичная Вторичная
4) Значения вторичной матрицы анализируются с позиций их сходства и различия. Существуют два основных способа такого анализа: кластерный и факторный. При кластерном анализе все значения, внесенные в матрицу, группируются попарно во всех возможных сочетаниях, и для каждой пары значений определяется степень математического сходства, например, Эвклидово расстояние (линейное расстояние между точками-таксонами в системе координат двух признаков). При факторном анализе компьютер одновременно рассматривает все таксоны в многомерном пространстве признаков, и выбирает 2 или 3 измерения-фактора (как правило, не соответствующие конкретным признакам), которые позволяют наиболее полно отобразить расстояния между таксонами в двух- или трехмерном пространстве соответственно. 5) Компьютер строит графическое представление данных, полученных в результате анализа сходств и различий. Данные кластерного анализа обычно представляют в виде кластерной диаграммы (дендрограммы, фенограммы) – древовидной схемы, в которой расстояние между таксонами выражено длиной связующих линий (рис.2). Результаты факторного анализа представляют в виде двух- или трехмерной системы координат, в которой таксоны указаны точками (рис.3). 6) Таксоны группируются друг с другом по принципу наибольшей близости в измерениях, предлагаемых кластерным и факторным анализами. Полученные группы, в свою очередь, объединяются в группы следующего порядка вплоть до соединения всех таксонов в единый супертаксон. Каждой из созданных групп присваивается определенный таксономический ранг. В результате создается иерархическая схема.
Достоинства Несомненным достоинством нумерического анализа является его объективность и научная строгость. Мощный математический аппарат позволяет точнейшим образом оценить многофакторное сходство и различие между организмами, выявить разрывы (хиатусы) между родственными группами особей, оценить степень корреляции между анализируемыми признаками. Это часто необходимо в исследованиях таксонов низкого ранга (родов, видов, разновидностей) и ископаемых форм. Когда возникают задачи такого рода, целесообразнее всего обращаться именно к нумерическому анализу. Недостатки Равенство признаков – крайне неоднозначное допущение. Вне зависимости от теоретического подхода (филогенетического, номотетического и т.п.) различные признаки будут иметь вовсе не одинаковую значимость для по- строения системы. Фенетические классификации, не учитывающие этот факт, часто оказываются теоретически бессодержательными3. Кроме того, нумерический анализ построен на оценивании сходства между организмами, и не способен выявить родства между ними, т.к. сходство зачастую имеет конвергентную природу. Несмотря на то, что кластерная диаграмма напоминает генеалогическое дерево, она не имеет никакого отношения к описанию хода эволюции и может найти крайне ограниченное применение в филогенетических исследованиях. А поскольку современная систематика опирается в первую очередь на филогенетический подход, применение в ней нумерического метода оказывается неактуальным, особенно в классификации высших таксонов. Кладистический метод Одним из наиболее широко распространенных методов классифицирования в настоящее время является кладистика, нашедшая применение в анализе как фенотипических, так и генотипических данных. Разработка кладистического метода связана с именем выдающегося немецкого биолога Вилли Хеннига (Hennig), который в 1950 г. опубликовал труд «Основные положения теории филогенетической систематики». В 1966 г. вышел в свет английский перевод этой книги, благодаря которому идеи В.Хеннига стали доступны мировой научной общественности. В том же 1966 г. была опубликована монография шведского энтомолога Л.Брундина (Brundin, 1966), который показал ценность методов, предложенных В.Хеннигом, для решения конкретных таксономических проблем. Так была заложена основа новой науки о реконструкции хода эволюции, которая получила название кладистика (от греч. clados - ветвь). В начале 1970-х гг. Г.Нельсон (Nelson), редактор журнала «Systematic Zoology», развернул на его страницах дискуссию о проблемах кладистики. Широкое обсуждение привело к повсеместному признанию молодой науки. C 1985 года в Нидерландах издается специализированный журнал «Cladistics». В течение 1980-1990-х гг. кладистическое исследование, так же как и фене-тическое, было полностью компьютеризировано, что существенно облегчило его проведение. В настоящее время наиболее распространенными ком- пьютерными программами кладистического анализа являются PAUP, NONA, McClade и некоторые другие. Принципы кладистики Будучи методологическим разделом самостоятельной науки, кладистиче-ский анализ построен на комплексе важных теоретических принципов, без детального знакомства с которыми трудно объективно оценить его возможности. Этих принципов четыре: •принцип синапоморфии; • принцип строгой монофилии; • принцип сестринских групп; • принцип парсимонии. Рассмотрим эти принципы подробнее. Принцип синапоморфии Все признаки организма кладисты оценивают с точки зрения их примитивности (плезиоморфности) и прогрессивности (апоморфности). Если несколько таксонов объединяет наличие одного апоморфного признака, то говорят о синапоморфии. Проявление же у двух таксонов общего плезио-морфного признака именуется синплезиоморфей. Наконец, наличие у таксона уникальных, ни у кого более не встречающихся апоморфных признаков называют аутапоморфией. Суть принципа синапоморфии заключается в том, что для построения филогенетической классификации имеют значения только синапоморфные признаки. Ни плезиоморфии, ни аутапоморфии не несут таксономической информации. Следует иметь в виду, что статус признака изменяется в зависимости от рассматриваемого иерархического уровня: апоморфный признак на определенном уровне рассмотрения оказывается синплезиоморфией. Например, пятипалые конечности человека, ящерицы и лягушки при сравнении с плавниками рыб являются синапоморфнфым признаком, и служат свидетельством того, что эти животные связаны общим происхождением. Однако у некоторых других наземных позвоночных задние конечности имеют не пять пальцев, а четыре (птицы), два (парнокопытные) или один (лошади). При выяснения филогенетических отношений между людьми, лягушками и лошадьми появляется возможность ошибочно противопоставить пятипалых людей и лягушек однопалым лошадям. Однако этого не произойдет, если следовать принципу синапоморфии. Дело в том, что однопалость является вторичной модификацией пятипалого состояния, т.е. апоморфна по отношению к нему. Следовательно, присутствие пяти пальцев у людей и лягушек на данном уровне рассмотрения является уже синплезиоморфией и не должно учитываться при построении классификации. Интересно оценить соотношение принципа синапоморфии с традиционными для эволюционизма понятиями гомологии и аналогии. Гомологией называ- ют сходство, обусловленное общим происхождением. При этом, считается, что наличие гомологичных органов является свидетельством принадлежности организмов к одному таксону. Однако, из приведенного выше примера следует, что гомологичными, т.е. обусловленными общим происхождением, являются и синапоморфные, и синплезиоморфные признаки, причем только первые из них действительно могут служить основанием для формирования группы. Итак, гомология в целом не является основанием для классификационных построений. Аналогия обычно понимается как сходство, не обусловленное общим происхождением. В кладистике такое сходство именуется гомоплазией и определяется как ситуация неоднократного возникновения признака в ходе эволюции.
Поиск по сайту: |