|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Происхождение и эволюция живых организмов
Наиболее сложным вопросом является вопрос о происхождении жизни. Дело в том, что до сих пор ученым не удалось экспериментально получить из неживых объектов живые организмы, не вовлекая в процесс какой-либо живой организм или его основной компонент. Тем не менее, имеются очень серьезные основания считать, что жизнь появилась на Земле примерно от 3,5 до 4 миллиардов лет тому назад в результате естественных химико-физических процессов, характеризовавших в то время состояние нашей планеты. Первое. Материальным субстратом всех биологических систем являются органические вещества, состоящие из элементов или химических групп, имеющихся на Земле – углерода, водорода, воды, азота, кислорода, серы, гидроксильной группы и многих других. Никаких экзотических, внеземных веществ в живых организмах на Земле не обнаружено. Второе. В настоящее время все биологические процессы так или иначе воспроизведены экспериментально: химические реакции, происходящие в живых организмах, процессы управления и взаимодействия с окружающей средой. Третье. Люди достаточно глубоко разобрались в механизме наследственности и изменчивости – главном процессе живой природы. Оказалось, что этот механизм вполне естественен, – он связан со строением и взаимодействием двух типов молекул – рибонуклеиновой и дезоксирибонуклеиновой, с помощью которых закодирована и транслируется наследственная информация. Четвертое. Археологические и эмбриональные изыскания убеждают нас в том, что формирование нынешней живой природы произошло путем постепенного усложнения структуры (развития) биологических организмов. Более того, оказалось, что это развитие соответствует законам существования диссипативных систем, установленным для неживой природы. Пятое. По всем своим признакам живые организмы соответствуют диссипативным управляемым системам. Они подчиняются всем законам физики и кибернетики. Шестое. Из всех планет Солнечной системы наилучшие условия для возникновения и существования жизни имеет Земля. В первую очередь это относится к диапазону температуры на поверхности Земли – от 200 до 350 Седьмое. Появившиеся в последние годы данные о том, что в определенных местах мирового океана существуют автономные биологические системы, не связанные с остальной живой природой [19]. На глубине до 3000 м на Срединно-Атлантическом хребте, там, где геосферные плиты раздвигаются выходящей из центральной части Земли магмой, находятся действующие вулканы, из которых вырывается расплавленная руда железа, меди и цинка, сопровождаемая черным дымом. И хотя вокруг такого вулкана, именуемого "черным курильщиком", безжизненное базальтовое дно с температурой всего 20С, сам он облеплен фантастическим оазисом с неизвестными формами жизни. В первую очередь это сероводородные бактерии, обитающие либо в виде колоний – толстых "матов", либо на лапках и жабрах существ, внешне напоминающих креветок. "Креветками" питаются крабы и голотурии – животные-цветы. Так на небольшом пространстве размером с трехэтажный дом в глубине океана существует автономный, оторванный от остального мира островок жизни. Восьмое. Если допустить, что жизнь возникла не на Земле, а где-либо в другом месте космического пространства, остается вопрос – а как она появилась там? И хотим мы того, или нет, мы вынуждены допустить, что таким местом может быть еще какая-нибудь планета с условиями, сходными с земными. Иначе жизнь, даже возникнув, не смогла бы устойчиво существовать, не говоря о распространении на другие планеты. Основные усилия учёных, изучающих проблему происхождения жизни на Земле, сосредотачиваются на поиске ответов на следующие вопросы: 1.Каковы основные условия, обеспечившие возникновение и устойчивое существование биологических систем? Попутно – как велика вероятность существования таких же условий на других космических объектах? 2.Насколько закономерно возникновение жизни даже при наличии тех условий, которые установлены при ответе на первый вопрос? 3.Каковы те основные физико-химические процессы, которые привели, в конечном счёте, к появлению основной структурной составляющей биологических систем – клетке? Приведём ответы на эти вопросы, содержащиеся в литературе последних лет [2,65, 66, 67, 68, 69]. Американский биохимик Л. Дж. Хендерсон (1878 – 1972г) в своём труде «Порядок природы» обосновал, что лишь наличие в неживой природе углерода С и воды Н2О обеспечивает возникновение жизни. Например, близкий к углероду по свойствам (и месту в таблице Менделеева) кремний образует значительно менее подвижный оксид SiO2 (песок), нежели абсолютно необходимый для формирования химических компонентов живого организма диоксид углерода СО2 (газ) [39]. Вода является главным фактором существования жизни. Поскольку жизнь – это сложная система химических реакций, – то наиболее приемлемой для нее средой является вода. Действительно, во-первых, она обладает самой высокой среди жидких неорганических веществ диэлектрической проницаемостью – в 81 раз больше, чем у вакуума. Это обеспечивает реакционную способность (т.е. способность вступать друг с другом в химические реакции) многочисленных веществ, растворённых или размешанных в ней. Во-вторых, удельная теплоёмкость воды выше, чем у большинства других веществ. Именно поэтому гидросфера Земли выступает в роли естественного стабилизатора температуры поверхности Земли, что очень важно для поддержания биологических процессов. Температура самой гидросферы – мирового океана – колеблется в узких пределах от +20С до 350С, что вполне приемлемо для химических реакций полимеризации органических веществ. Еще одно, третье важнейшее свойство воды – при замерзании её плотность уменьшается. Поэтому водоёмы при температуре ниже нуля по Цельсию оказываются естественным образом покрыты теплоизоляционной оболочкой. В-четвёртых, вода обладает более высокой температурой и более широким диапазоном температур жидкой фазы, нежели другие естественные аналоги: аммиак (NH3) – -780 ... -330С; двуокись углерода (СО2) – -570 ... -790С; метан (СН4) – -1840 ... -1620С; цианистый водород (HCN) – -140 ... 260С. Это также очень важное обстоятельство, так как обеспечивает наиболее высокую скорость протекания химических реакций между органическими полимерами. В-пятых, вода обладает большим, чем другие жидкости, поверхностным натяжением и поэтому легко поднимается по капиллярам почвы и тканей растений. В-шестых, наличие водородных связей между молекулами воды Н2О приводит к созданию полимолекул (Н2О)n, структура которых зависит от начальных условий. Таким образом, вода обладает исходными признаками памяти, что, по-видимому, также сыграло важную роль на первом этапе возникновения жизни. Согласно [66,67], гидросфера Земли появилась не менее, чем 3.8...4.0 млрд. лет тому назад. Надо полагать, что именно тогда начался второй подэтап первого этапа биологической эволюции – см. ниже. Следующим фактором, обеспечившим возникновение жизни на Земле, является её масса и плотность. С одной стороны их величина обеспечивает удержание довольно толстого слоя атмосферы (например, у Марса и Луны, масса которых значительно меньше земной, атмосфера практически отсутствует). С другой стороны, эта величина не так велика, как, скажем, у Юпитера или Сатурна, что существенно уменьшает силовую нагрузку на живые организмы. Кроме этого, повышение давления смещает температурный диапазон жидкой фазы воды в область более высоких температур. Ещё одним важнейшим фактором, обеспечившим возникновение жизни, является масса Солнца. Как было сказано в главе четвёртой, у звезды именно такой массы, как Солнце, длительность первой, основной фазы эволюции составляет 7...10 млрд. лет. Это обеспечивает стабильный температурный и радиационный режим поверхности Земли на протяжении срока, необходимого для превращения химической (абиогенной) эволюции в биологическую, т.е. нескольких миллиардов лет. Все перечисленные условия являются необходимыми для зарождения жизни. Какова вероятность появления планеты с условиями, схожими с земными? Расчёты, проведённые рядом специалистов [2,35], показывают, что эта вероятность составляет миллиардные доли процента. Однако, учитывая, что общее число звёздных систем составляет 1021, нетрудно заключить, что во Вселенной могут находиться миллиарды планет, аналогичных Земле. Следующий вопрос – о неизбежности возникновения жизни при наличии перечисленных выше условий – получает всё большее экспериментальное подтверждение. Во-первых, в космическом пространстве (газовых облаках, метеоритах и кометах) обнаружены достаточно сложные органические соединения [67] – формальдегид Н2СО, муравьиная кислота НСООН, цианоацетилен НС3N, формамид НСОNН3. Предполагать, что эти вещества затем оказались в составе поверхностного слоя Земли и положили начало химической эволюции, было бы неправомерным. Дело в том, что начальная температура этого слоя, которая продержалась несколько сот млн. лет, составляла 800...9000К. При такой температуре эти вещества должны были распасться. Однако сам факт появления этих веществ на космических объектах свидетельствует о естественности их синтеза. Во-вторых, многочисленные лабораторные исследования [67] убеждают в том, что в условиях, существовавших на начальном этапе эволюции Земли, возникновение органических соединений, составивших материальную основу живых организмов, было неизбежным. Судя по всему, из пород, сформировавших Землю, в атмосферу выделялся метан СН4, аммиак NН3, азот N2, оксид углерода СО2 и СО, сероводород Н2S и пары воды Н2О. Основным источником энергии, воздействовавшим на эту атмосферу, было солнечное излучение. Выше (см.§3.4) указывалось, что на 1м2 земной поверхности попадает 1010 Дж солнечной радиации (по всему частотному спектру – от инфракрасного до g- лучей) в год. Эта энергия соответствует [49] потоку информации 6´1014 Вт / 0К на всю поверхность Земли или 1023 байт на 1 м2 ежесекундно. Энергия всех других источников (радиоактивный распад, вулканические процессы, разряды) меньше солнечной в 104...106 раз. Ни в коей мере не исключая роль всех перечисленных источников, исследователи основное предпочтение отдают фотопроцессам, обусловленным солнечной энергией. При этом весьма существенную роль играют каталитические свойства некоторых соединений кислорода, кремния, алюминия, кальция, магния и титана, входящих в состав земной коры. В соответствии с имеющимися археологическими данными возникновение и развитие жизни на Земле можно разбить на три главных этапа. Первый этап занимает период от 4,5 до 3,5 млрд. лет тому назад, т.е. от момента возникновения Земли до появления первых одноклеточных живых существ. Этот период "абиогенного синтеза" [76] характеризуется появлением предбиологических соединений и образованием из мирового океана, покрывавшего всю поверхность Земли, "первичного бульона". Второй этап длился наиболее долго - от 3,5 до 0,8 млрд. лет тому назад. Это период формирования и бурного размножения одноклеточных в мировом океане. По сути дела в этот период жизнь прошла путь от продукта неживой природы Земли к активному формированию ее структуры. Начиная с этого периода, строение верхних слоев литосферы, состав атмосферы и гидросферы во многом определяется живыми организмами, в том числе одноклеточными. В этот период окончательно сформировалась сложнейшая структура клеток, предопределившая дальнейшее развитие жизни. Третий этап начался 600 - 1000 миллионов лет назад и длится поныне. Это период возникновения и бурного развития многоклеточных живых организмов – царства растений, царства грибов и царства животных. В свою очередь, первый этап абиогенного синтеза или химической эволюции можно разбить на два подэтапа. Оба подэтапа отличаются друг от друга тем обстоятельством, что в течение первых 500¸600 млн.лет существования Земли температура на её поверхности была выше 1000С и лишь 3.9¸4.0 млрд.лет тому назад она остыла настолько, что появилась гидросфера. Именно этот рубеж и является разделительным для обоих подэтапов. На первом подэтапе температура атмосферы Земли, её химический состав, жёсткое ультрафиолетовое облучение Солнца и активная вулканическая деятельность обусловили возникновение базовых органических веществ. Атмосфера Земли мало отличалась от атмосфер других планет Солнечной системы и состояла, в основном, из простейших соединений углерода, водорода, азота между собой и с кислородом: NH3, CH4, H2O, HCN, NO(NO2,…), CO и СО2. К этим компонентам добавлялись вырабатываемые извержениями вулканов P, S, Cl и ряд других элементов. Многочисленные лабораторные исследования последних десятилетий показали, что под воздействием ультрафиолетового излучения и электрических разрядов, сопровождавших извержения вулканов, возникают аминокислотные мономеры (пептиды), нуклеотиды, сахариды и липиды. Неизбежность этих химических реакций подтверждает обнаружение базовых органических веществ в ряде метеоритов и на других планетах Солнечной системы. В таблице 5.2 приводятся результаты одного из первых опытов по синтезу аминокислот при воздействии ультрафиолетового облучения и электрических разрядов на смесь,имитирующую первичную атмосферу, и обнаруженные в одном из метеоритов остатки аминокислот. Таблица 5.2 Выход природных аминокислот при имитации предбиологического синтеза: опыт Миллера (пропускание электрического разряда через смесь CH4, N2, и Н2О) и в метеорите Мергинсона
Процесс синтеза сложных органических молекул протекал в такой последовательности [81]. Сначала, поглощая кванты длинноволнового излучения, молекулы NH3, H2O, CH4, N2 и СО2 распадаются соответственно на (NН2+Н),(ОН+Н), (СН3+Н) и (СН2+Н2), (N+N), (СО+О). Затем из этих промежуточных продуктов образуются простейшие органические соединения – формальдегид, цианистый водород, формамид. Затем, уже после образования водоёмов, начали синтезироваться более сложные соединения – липиды, сахариды, порфирины, аминокислоты, нуклеотиды. И, наконец, уже в водоёмах, по-прежнему под воздействием солнечной радиации, стал происходить процесс полимеризации этих соединений. Таким образом, химический состав атмосферы и поверхности Земли, а также интенсивное воздействие солнечной радиации привели 3,5...4 млрд. лет тому назад к тому, что на поверхности Земли возникли водоёмы, насыщенные органическими молекулами, которые находились в непрерывном химическом взаимодействии под влиянием солнечной радиации. Следовательно, первые живые организмы были продуктами интенсивной химической эволюции. Эта эволюция привела не только к подавляющему доминированию полипептидов - катализаторов, но и к существенному усложнению каталитических процессов. Среди многочисленных вариантов сложных взаимозависимых химических реакций появились циклические. Циклическая последовательность химических реакций – это такая, при которой периодически возникает одна и та же концентрация одних и тех же веществ. Примером таких циклов является реакция Белоусова – Жаботинского, описанная в[16]. Следует, однако, отметить, что реакция Белоусова - Жаботинского постепенно угасает, так как концентрация её исходных веществ с каждым циклом уменьшается. Её можно поддерживать сколь угодно долго, если постоянно подавать в раствор исходные компоненты и убирать шлаки в постоянно увеличивающемся количестве. Именно такие циклические реакции возникли на втором подэтапе первого этапа в «первичном бульоне» согласно гипотезе М. Эйгена и П. Шустера [68]. Основными компонентами этих реакций были полипептиды и полинуклеотиды. Роль первых – инициировать вполне определённые реакции полимеризации аминокислот и нуклеотидов. Роль вторых – «координировать» структуру образовавших их аминокислотных полимеров, и катализировать формирование дубликатов этих последних. Второй подэтап, наступивший после снижения температуры поверхности Земли ниже 1000С и образования Мирового океана, явился, судя по всему, инициирующим в процессе возникновения жизни. Его можно разбить на следующие стадии: 1.Полимеризация пептидов и нуклеотидов, преобразование Мирового океана в коллоидный раствор этих полимеров – «первичный бульон». 2.Дальнейшее снижение температуры гидросферы и возникновение «коацерватов». 3.Компартментация (покрытие полупрозрачными оболочками) коацерватов и возникновение «гиперциклов». 4.Эволюция гиперциклов, обусловленная их конкуренцией, возникновение процессов трансляции и репликации – превращение гиперциклов в «пробионты». Рассмотрим эти стадии подробнее, а заодно раскроем содержание тех терминов, которыми они именуются. Первая стадия – возникновение «первичного бульона» - длилась не менее 100¸200 млн.лет и характеризовалась относительно высокой температурой воды – не менее 500С. Эта температура способствовала полимеризации пептидов и нуклеотидов, последовательности которых носили, естественно, случайный, стохастический характер. Это не исключало их взаимное каталитическое влияние, поэтому в коллоидном растворе их в воде возникла химическая эволюция, о которой говорилось в § 3.5, и которая вела к увеличению концентрации химически активных полимеров и постепенному исчезновению нейтральных. Одновременно и в атмосфере и в гидросфере продолжалось, всё более увеличиваясь в производительности, производство исходных, базовых органических веществ, о которых говорилось выше. Вторая стадия – стадия возникновения «коацерватов» – характеризуется дальнейшим снижением температуры воды до стационарного уровня, который с небольшими колебаниями сохранился до нашего времени. При такой температуре – + 2¸350С – в коллоидном растворе полипептидов образуются коацерватные капли или просто коацерваты – клубки (глобулы) полипептидов, о химических причинах возникновения которых сказано в §5.6. Вокруг каждой такой капли формировалась оболочка из молекул воды. Эта оболочка частично изолировала коацерват от окружающей среды, создавая исходные предпосылки для компартментации. Однако эта оболочка не мешала полимерным молекулам коацервата присоединять к себе аминокислотные мономеры из окружающей среды. Такой способ обмена веществ именуется гетеротрофным [28,29]. Капля естественным образом росла и, достигнув определённых размеров, разваливалась на несколько «дочерних» капель в силу причин, изложенных в §5.2. Эти новые капли вовсе не повторяли процессы «родительской», а «жили» новой, совсем непохожей «жизнью». Единственным системным процессом в этом потоке коацерватов была химическая эволюция, т.е. увеличение концентрации химически активных полипептидов в них. Длилась стадия также, по-видимому, несколько десятков миллионов лет. Третья стадия – возникновение «гиперциклов» - наступила вследствие продолжающейся химической эволюции. В результате происходило не только увеличение химической активности полипептидов, но и появление более системно сложных каталитических процессов. В частности, появились коацерваты с замкнутыми циклами инициирующих друг друга химических реакций. Особую роль играли в них вплетённые в глобулы полипептидов полинуклеотиды. Такое переплетение представляется естественным, учитывая случайный характер формирования химического состава коацерватов. Полинуклеотиды в силу своих свойств кодировали последовательности аминокислот входящих в каплю полипептидов и благодаря этому вносили определённую упорядоченность в замкнутый цикл химических реакций. Такая последовательность взаимозависимых полипептидов , полинуклеотидов и связывающих их химических реакций полимеризации именуется гиперциклом [6, 49, 68]. Первоначально гиперциклы формировались полипептидами, образованными всего двумя наиболее примитивными аминокислотами – глицином( радикал Rсостоит из атома водорода H) и аланином(радикал состоит из метиловой группы СН3), и нуклеиновыми кислотами, образованными всего двумя нуклеотидами – Ги Ц. Интересно, что глицину соответствует комбинации ГГГ и ГГЦ , а аланину ГЦГ и ГЦЦ. Поэтому можно предположить, что полипептиды, состоящие из набора глицина и аланина, катализировали полимеры, состоящие из нуклеотидов Ги Ц, причем в такой последовательности, которая соответствовала Глии Алав исходных молекулах – катализаторах. Важнейшим событием эволюции гиперциклов стало возникновение репликации входящих в них полинуклеотидов. Судя по всему, катализаторами такой репликации (синтеза пары молекул – дублёров) выступали первоначально сами исходные полинуклеотиды по схеме сдваивания цепи в транспортных РНК(тРНК) – [49]. Из всех известных в настоящее время нуклеиновых кислот только тРНК (транспортные РНК) вступают непосредственно в химическую реакцию с аминокислотами. Поэтому очевидно, что молекулами НК, которые явились активными участниками абиогенной эволюции, были предшественники тРНК [49, 68]. Основные структурные особенности этих молекул: 1)образование петель, которые замкнуты параллельными цепочками комплементарных нуклеотидов (например, есть участок, в котором оба участка соединены
Т А Т Т Г Г А Ц Ц Т А А Ц Ц Т Г(5.11)
водородными связями);2) наличие активного конца, который способен соединяться с одной из 20 биологических аминокислот; 3) наличие петли, содержащей в своей вершине (кольце) последовательность трёх нуклеотидов, образующих антикодон той самой аминокислоты, которая присоединяется к свободному концу молекулы – игле (например, триплет
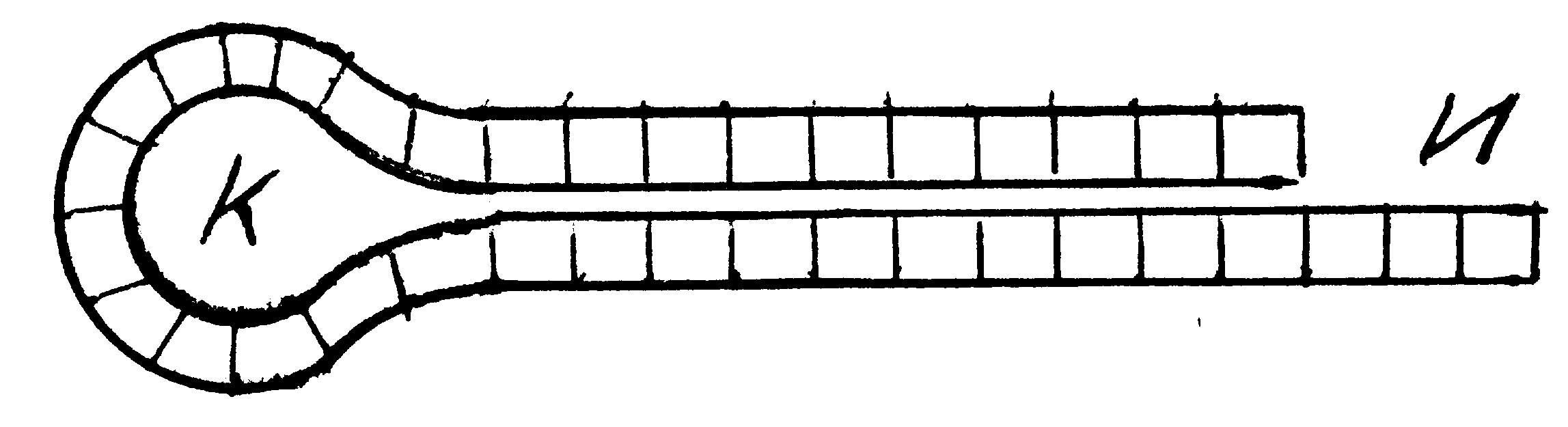
Ц Ц Г для тРНК, соединяющейся с глицином); 4) высокая подвижность молекулы внутри полипептидного геля. Всё это делает молекулы типа транспортной РНК наиболее приемлемыми для формирования каталитического взаимодействия между полимерами аминокислот и нуклеотидов. С учётом сказанного общая картина перерастания химической эволюции в биологическую представляется в рамках теории гиперциклов следующим образом. Вместе с коацерватными каплями, состоящими в основном из полипептидов, в первичной гидросфере Земли возникли и эволюционировали полинуклеотиды (нуклеиновые кислоты - НК), состоящие в основном из гуанина и цитозина. Если длина полимерной цепочки НК состояла из менее, чем 100 узлов, последняя быстро распадалась под воздействием гидролиза. Следовательно, все устойчиво существовавшие молекулы НК содержали не менее 100 нуклеотидов. Эти молекулы могли путём притягивания к каждому своему узлу нуклеотида – двойника (например, к гуанину цитозина) сформировывать свои антиподы (например,антиподом Ц-Ц-Ц-Г-Ц-Г-Г-Ц служит молекула Г-Г-Г-Ц-Г-Ц-Ц-Г).Таким образом, мировой океан наполнился нуклеиновыми кислотами с одинаковым или зеркально-противоположным строением. Часть этих молекул деформировалась в петли, состоящие из круглого кольца К и иглы И:
(5.12)
Многие вершины петель подсоединялись к антикодоновым участкам линейных полипептидов, игравшим роль памяти:
(5.13)
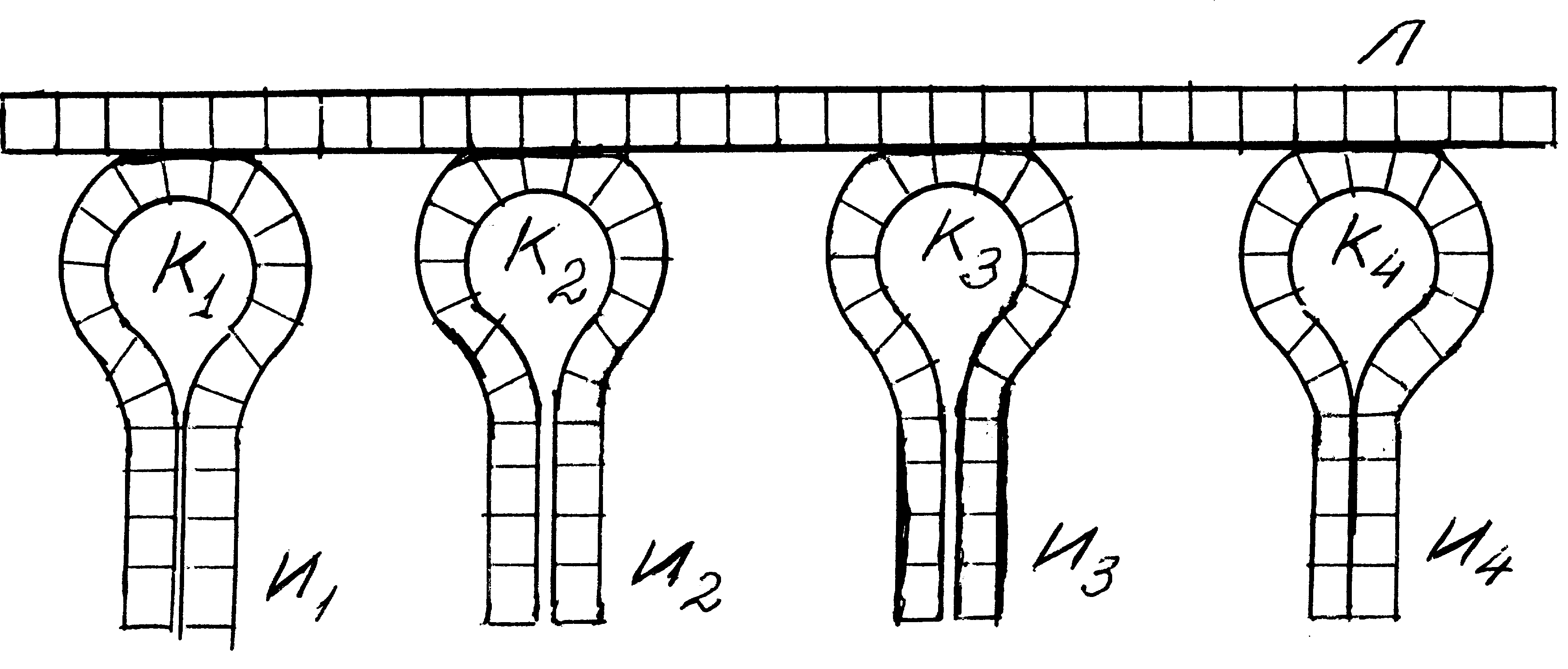
Таким образом, на определённой фазе химической эволюции мировой океан оказался заполнен коацерватами из весьма химически активных полипептидов, достаточно однотипных нуклеиновых цепочек и их антиподов (±НК), конструкций из этих нуклеиновых цепочек и приклеившихся к ним петлеобразных НК. Когда эти конструкции оказывались вплетёнными в коацерваты, они начинали «вылавливать» в окружающем «бульоне» иглами И молекулы Гли и Ала и формировать из них полимерные цепочки. Ясно, что путём отбора последовательности этих пептидов стали образовывать ферменты, катализировавших, во-первых, соединение петлеобразных НК с аминокислотами и, во-вторых, самополимеризацию, т.е. «склеивание» в полимерную цепь молекул Гли и Ала, находившихся на «иглах» соответствующих НК.Все другие комбинации коацерватов с нуклеиновыми кислотами распадались. Из описанных выше конгломератов образовались первые гиперциклы. Условно их можно изобразить так [68]:
(5.14)
где I – совокупность полимеров нуклеотидов (информационные интермедиаты – участники циклических реакций), Е – совокупность полипептидов – катализаторов (силовые интермедиаты). Данные гиперциклы обладали главным свойством биологических систем – они делились. Благодаря тому, что линейные полинуклеотиды Л (5.13) дублируются, «дочерние» гиперциклы повторяли процессы в «родительском». Таким образом, после распада родительского коацервата в одной из дочерних капель под воздействием уже сформировавшегося полимера нуклеотидов Ги Цобразовывался полипептид, повторивший родительский. Далее процесс циклически повторялся. (Отсюда и название – гиперцикл). Так определились следующие фазы стадии гиперциклов: 1. Свободное существование в первичном бульоне коацерватов полипептидов и нуклеиновых кислот. 2. Возникновение комплементарных цепочек, петель (5.12) и конгломератов (5.13). 3. Зарождение кодирования и трансляции по схеме (5.13), а также репликации. 4. Гиперциклическая фиксация кода с ГЦ – рамкой (по схеме 15.14), кодирование Гли, Ала, Асп и Вал. 5. Эволюция гиперциклического кода, репликации, синтеза белка, включение в гиперцикл гидрофобных аминокислот, компартментация. 6. Возникновение полностью изолированных гиперциклов, адаптированных ферментов репликации и трансляции, эволюция функций метаболизма и контроля, появление оперонной структуры, превращение НК в РНК современных вирусов. 7. Преобразование гиперцикла в протоклетку, появление геномов – ДНК, усложнение системы ферментов, системы контроля; кодирования и считывания наследственной информации. Именно на этом этапе химической эволюции возникло новое природное явление – наличие целесообразности процесса. Два соседних гиперцикла, если они отличались друг от друга, либо вступали в конкурентную борьбу и гибли, либо начинали «сотрудничать» с целью выживания. В этом последнем случае возникает более сложный гиперцикл:
(5.15)
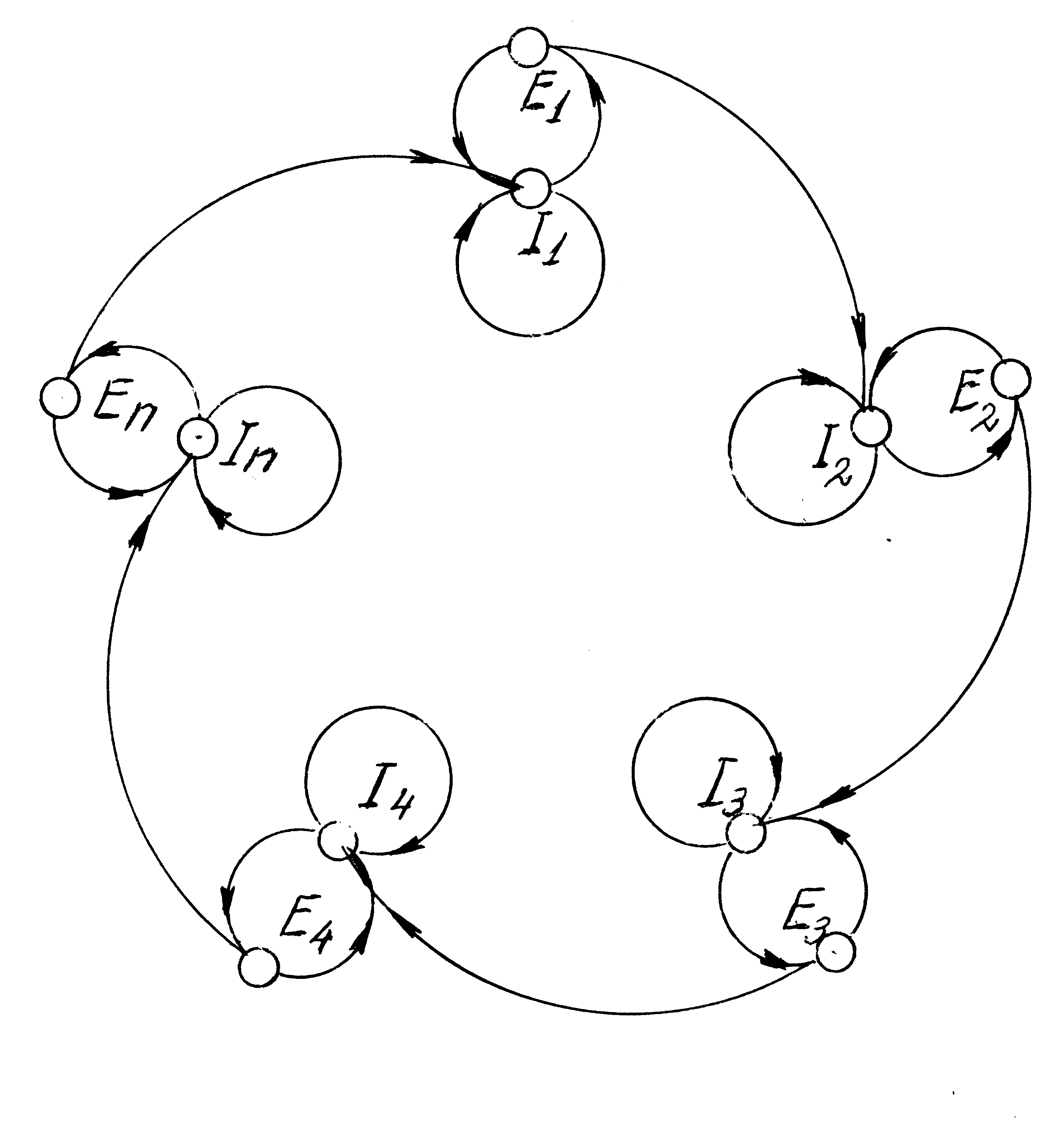
Ферменты Е1 интенсифицировали активность I2, а Е2 – I1. Так возникал новый, ещё более сложный гиперцикл. Линейные полимеры I1 и I2соединялись, совмещая в себе «гены» Е1и Е2. Далее в процесс постоянного видоизменения химического состава гиперциклов включился процесс мутации нуклеотидов – некоторые пурины Гпревращались в А, а пиримидины Ц – в У.Новые информационные молекулы включали в замкнутые каталитические цепи новые аминокислоты. Возникшие гиперциклы, также как уже существовавшие, в результате конкуренции либо гибли, либо образовывали друг с другом всё более сложные гиперциклы:
(5.16)
Появление дочерних коацерватов – гиперциклов, в значительной степени повторявших по структуре и процессам родительские, кардинальным образом изменило ситуацию в природе. Дело в том, что произошло включение механизма мальтусовского процесса [2] накопления в геометрической прогрессии одинаковых объектов. Действительно, пусть такая предклетка делится два раза в час, тогда через сутки таких коацерватов будет 248»1016, т.е. десять миллионов миллиардов. А сколько их будет через неделю, месяц, год… При этом в силу гетеротрофного характера питания модернизированных коацерватов эти последние «поедали» все питательные вещества окружающего океана. В конечном счёте, вследствие этого должна была возникнуть конкурентная борьба между гиперциклами. Это обстоятельство и привело к появлению нового механизма эволюции – естественному отбору. Ясно, что из двух конкурирующих гиперциклов сохраняется тот, у которого совершеннее система запоминания достигнутого порядка. И, наконец, четвёртая стадия первого этапа – стадия возникновения пробионтов. Появившиеся в результате эволюции гиперциклов пробионты представляли собой объекты, находившиеся на границе живого с неживым. У них, в отличие от гиперциклов была значительно более чётко выраженная компартментация, т.е. обособление внутренней среды от внешней с помощью оболочки, отдалённо соответствующей мембране и включавшей в себя липиды. Полинуклеотиды постепенно преобразовались в пред-РНК или ядерные РНК, которые освоили функции трансляции (преобразование кода последовательности нуклеотидов в последовательность аминокислот в полипептидах) и репликации (деление на две дочерние молекулы, полностью повторяющие информацию исходной). Однако эти пробионты ещё не были клетками, так как в них отсутствовали многие структурные органеллы, обеспечивавшие метаболизм и жизнедеятельность. Так завершился этап предбиологической (абиогенной) эволюции. Второй этап становления жизни на Земле – этап возникновения и развития одноклеточных организмов – также условно можно разбить на два подэтапа: Подэтап прокариотов – 3,5 ¸2,0 млрд.лет тому назад, и подэтап эукариотов – 2.0¸0.8 млрд.лет тому назад. В свою очередь, первый подэтап подразделяется на три стадии: 1.Стадия возникновения и эволюции гетеротрофных анаэробных прокариот – 3.5¸3.0 млрд.лет. 2.Стадия образования архебактерий – 3.0¸2.5 млрд.лет. 3.Стадия возникновения биологического механизма образования пептидов и нуклеотидов и появления автотрофного метаболизма – 2.5¸2.0 млрд.лет. Второй подэтап также подразделяется на ряд стадий: 1.Образование эукариотов. 2.Возникновение механизма целенаправленных мутаций. 3.Возникновение полового размножения. 4.Разделение эукариот на авто- и гетеротрофных. Возникновение аэробного механизма окисления. Представляется, что этот этап является решающим в длительном процессе «сотворения» живой природы. Трудности воспроизведения в лабораторных условиях процесса зарождения жизни связаны главным образом с тем, что этот процесс не одного дня, месяца, года, столетия, а сотен миллионов и миллиардов лет. Первые живые организмы – анаэробные клетки-прокариоты гетеротрофного типа – появились, судя по ряду достаточно доказательных археологических данных, примерно 3,5 млрд. лет тому назад как итог эволюции пробионтов. У них была чётко выраженная мембранная оболочка, современная «азбука» кодирования, считывания и репликации наследственной информации и начальный механизм внутреннего регулирования химических процессов. Носителем наследственной информации этих клеток были молекулы РНК, содержащие всего несколько тысяч нуклеиновых узлов. Питались эти клетки продуктами абиогенного синтеза, которым был наполнен Мировой океан. О том, насколько концентрированно были заполнены воды Мирового океана этими продуктами, можно судить по минеральным залежам этого океана – нефтеносным слоям. Тем не менее, быстрое размножение клеток позволяло им в кратчайшие сроки истощать окружающие воды и заменять растворённые в них органические вещества своими потомками. Условия для мутаций этих клеток были достаточно благоприятными: не было устранено интенсивное облучение поверхности Земли ультрафиолетовыми и другими электромагнитными волнами, продолжалась бурная вулканическая деятельность с выделением активных химических веществ и электрическими разрядами. Однако подавляющее большинство этих мутаций только нарушало сложившийся порядок взаимодействия полипептидов и полинуклеотидов в прокариотах. Лишь те мутации, которые увеличивали записанную в наследственных РНК информацию (а значит, увеличивали саму молекулу РНК), позволяли осуществляться процессу самосовершенствования клеток. Эти мутации осуществлялись, по-видимому, двумя естественными путями. Во-первых, в клетках, в дополнение к уже запомнившемуся циклу обмена информацией между ферментами и РНК-памятью, возникал ещё один (по крайней мере) гиперцикл, который привёл на ранней стадии эволюции к появлению основной обратной связи . Этот дополнительный контур обратной связи осуществлял функции второго каскада сложной системы самоуправления. Во-вторых, клетки поглощали из окружающей среды не только изолированные друг от друга мономеры аминокислот и нуклеотидов, но и их полимеры – остатки погибших клеток. Не исключено, что вместе с этими поглощениями они получали новую информацию, которую добавляли к своей. По-видимому, так появились и предки вирусов – изолированных молекул РНК и ДНК, способных функционировать только внутри клетки. Однако не все возникавшие таким образом изменения обязательно повышали устойчивость сохранения клетки. Далее действовал механизм естественного отбора. Совершенно очевидно, что только те мутационные изменения РНК, которые в процессе жизнедеятельности одной клетки включались в цикл взаимосвязи вместе с возникшими ферментами, могли обеспечить дальнейшую эволюцию клеток. Итак, ещё одним отличительным признаком, которым обладали прокариоты по сравнению с пробионтами, было наличие дополнительного каскада обратной связи, обеспечившего целенаправленные (конструктивные) мутации. Очевидно, именно это обстоятельство привело к тому, что появились археи (или архебактерии [67]) – клетки, приспособленные жить в экстремальных, не очень-то благоприятных для жизнедеятельности условиях. Археи – весьма простые организмы, отличающиеся от прокариотов системой синтеза белка (рибосомальной системой), наружной мембраной, биохимическими процессами. Впрочем, некоторые исследователи считают, что археи и прокариоты произошли от общего предка – прогенота. Главная особенность архей – способность к жизнедеятельности в весьма необычных для жизни условиях. Так, например, одна из разновидностей архей – кренархеоты - существуют при температурах выше 800С вблизи «чёрных курильщиков» (см. §6.1), используя для жизни (окислительно-восстановительных реакций) находящуюся в вулканическом газе серу. Другая разновидность – эвриархеоты – обнаружены не только вблизи «чёрных курильщиков», но и в прямо противоположных по температурным условиям зонах – в арктической тундре и Антарктиде. Энергию окислительно-восстановительных реакций они берут из остатков органической среды (например, соли муравьиной или уксусной кислоты, метиловый спирт и метиламины). Некоторые из них «дышат» углекислым газом, выделяя метан: 4Н2 + СО2 = СН4 + 2Н2О. Эти простейшие организмы часто являются симбионтами (т.е. участниками симбиоза – биологического содружества) некоторых животных, синтезируя, например, витамин В12. Ещё одна группа архей – галофильная – способна к жизнедеятельности только в насыщенных солевых растворах. В частности, они образуют своеобразную фауну Мёртвого моря на Ближнем Востоке, соляных озёр Африки, Америки и даже юга России. Особую группу архей составляют кислотолюбивые. Они развиваются в такой кислотной среде, которая может вызвать, например, ожог кожи человека. Совершенно очевидно, что археи появились как один из способов приспособления быстро размножавшихся простейших живых организмов к условиям, непривычным для исходных биологических систем. Благодаря такому приспособлению они избавились от конфликта с другими организмами, и поэтому у них пропала необходимость к структурному совершенствованию. Но самое главное – с появлением археев на Земле появился ещё один источник органических веществ, необходимых для жизнедеятельности биологических систем. До них единственным источником таких веществ был абиогенный синтез. В дальнейшем из архей (предположительно) появились бактерии, способные перерабатывать, используя солнечный свет, углекислый газ СО2 и воду Н2О в органические компоненты – предки хлоропластов растений. Это привело, в конечном счёте, к тому, что в настоящее время основные органические вещества – материальная основа живых организмов – вырабатываются из неорганических веществ самими биологическими системами – растениями, цианобактериями и т.п. Ими синтезируется ежегодно 1011 тонн органического вещества [67]. Так наступила вторая стадия первого подэтапа эволюции одноклеточных – стадия появления архей. Именно у этих, также весьма примитивных клеток, появился механизм автотрофного питания, т.е. биологического формирования пептидов и нуклеотидов. Одновременно продолжался процесс совершенствования прокариотов. Носителем наследственной информации стала молекула ДНК, что сразу резко повысило устойчивость клеток к разрушительным воздействиям внешней среды. Так сформировался основной ствол эволюции прокариотов, связанный с объединением их наследственных признаков (симбиозе), и побочные ветви, позволившие выжить вытесненным из основного массива Мирового океана бактериям и приспособиться к экстремальным для биохимических процессов условием, – многочисленные ветви археев. Размножение архей постепенно привело к тому, что абиогенный синтез органических веществ всё более сменялся на биологический. Третья стадия, возникшая благодаря произошедшим изменениям, характеризуется, во-первых, тем, что механизм биологического синтеза аминокислот и нуклеотидов практически вытеснил абиогенный, во-вторых, живые организмы всё более начали влиять на формирования наружных слоёв Земли – атмосферы, коры и гидросферы, и, в-третьих, ускорением самосовершенствования самих клеток. На этой стадии возник или, во всяком случае, занял подобающее место ещё один механизм усовершенствования клеток – образование симбионтов. Две приблизившиеся друг к другу клетки вместо того, чтобы вступать в единоборство, начинают «сотрудничать» для совместного выживания. Так возникли митохондрии, хлоропласты, органы движения клеток – жгутики. На этой же стадии появился новый класс ферментов – пигменты, использующие энергию световых лучей для синтеза органических веществ. На следующем подэтапе «царями» живой природы стали эукариоты. В среднем эукариоты больше прокариотов в несколько сот раз. Судя по всему, главным фактором, приведшим к превращению прокариот в эукариоты, был механизм образования симбионтов. Сотрудничавшие между собой прокариоты и археи, объединившись, превратились в органеллы более сложного объекта. Одновременно увеличивалось разнообразие различных видов живых организмов. Постоянная конкурентная борьба между ними вынуждала усовершенствовать их механизм наследственности. При этом далеко не всегда возникали более сложные клетки. В подавляющем большинстве случаев популяциям удавалось приспособиться к условиям существования и без дальнейшего усложнения их организма. Однако ни на минуту не ослабевавшая борьба за выживание приводила к возникновению внутри некоторых видов более сложных разновидностей, которые давали побеги дальнейших эволюционных ветвей и основного ствола развития. Ниже мы будем отслеживать процессы, связанные именно с этим стволом развития, не забывая о том, что появлявшиеся в нём виды образовывали попутно и боковые ветви эволюции. Очень важным фактором развития эукариот было усовершенствование механизма приспособления к условиям окружающей среды – механизма наследственности. Как уже указывалось выше, мутации, вызвавшие непосредственное изменение структуры молекулы ДНК под воздействием физико-химических процессов в окружающей среде, не могли служить основой конструктивного «творчества» природы. Лишь те мутации (регенеративные), которые были связаны с появлением новых замкнутых обратных связей в клетке между полипептидами и нуклеотидами, служили основой для появления более сложных её свойств. Важным «изобретением» природы стало появление в ДНК подвижных генов – «транспозонов» [59, 62, 68]. Естественная творческая лаборатория пополнилась ещё одним эффективным инструментом, который привёл к существенному ускорению прогресса. Ещё более усовершенствовало механизм изменчивости появление полового размножения. Главное достоинство этого способа – постоянный обмен генетическим материалом между всеми особями популяции. Действительно, молекулы ДНК клетки, относящиеся, например, к 30-му поколению своей популяции, несут в себе информацию, полученную от 230 »109 (миллиарда) особей. Конечно, среди её предков могут быть «родственники», тогда общее число первых будет меньше. Зато появляется возможность культивировать более полезные для условий среды признаки. Сложный обмен «родственными» генами превращает всю популяцию в «процессор» с очень сложной структурой взаимной передачи информации. Не удивительно, что в этом «процессоре» появились сначала простые, а потом всё более усложняющиеся эвристические алгоритмы. И, наконец, завершающая стадия этапа одноклеточных ознаменовалась окончательным разделением эукариот на гетеротрофных и автотрофных. Последние, освоив механизм хлорофильного синтеза белка и других биопродуктов, положили начало царству растений, а первые – царствам грибов и животных. Так примерно 600 – 800 млн.лет тому назад эукариоты достигли такой степени развития, что могли соперничать между собой за выживание, лишь создавая многоклеточные колонии. К этому времени и естественная среда для развития живых организмов (также в результате воздействия их предшествующей эволюции) существенно изменилась и способствовала жизнедеятельности многоклеточных. В первую очередь, это связано с появлением чистого кислорода в воздухе, обеспечившего аэробное питание и создание защитного озонного слоя. Для подтверждения сказанного сошлёмся на [67], согласно которой современная палеонтология интерпретирует геологические данные в виде следующей хронологической последовательности главных событий на 1 и 2 этапах эволюции жизни на земле: 4 млрд. лет назад – образование океанов и материков (гидросферы); 4.2 ... 3.2 млрд. лет – химическая эволюция (абиогенный синтез), завершившаяся созданием анаэробных бактерий; 3.2...2.9 млрд. лет – возникновение фотосинтезирующих бактерий; 3...2.5 млрд. лет – увеличение разнообразия анаэробных прокариотов; 2.5 ... 2.1 млрд. лет – развитие аэробного синтеза; 2.1 ... 1.9 млрд. лет – появление чистого кислорода в атмосфере, развитие аэробного дыхания, вымирание некоторых анаэробов, возникновение эукариотов; 1.9...1 млрд. лет – увеличение разнообразия аэробных прокариотов, увеличение разнообразия эукариотов, появление и развитие полового размножения; 1 ... 0.8 млрд. лет – возникновение мегаскопических эукариотов и увеличение их разнообразия, появление простейших многоклеточных организмов; 0.8 ... 0.5 млрд. лет – развитие твёрдых частей у водорослей и Метаzoa. Надо сказать, что некоторые формы живых организмов выходят за пределы приведенных временных рамок. Так, согласно [43], низшие грибы (заведомо эукариоты) найдены в породах, возраст которых около 2.2 млрд. лет тому назад, а следы первых многоклеточных животных находят в слоях, возраст которых около 1.4 млрд. лет. Однако это лишний раз подтверждает тот факт, что эволюция биосферы Земли – это не строго детерминированный, а стохастический процесс, основные закономерности которого определяются в масштабах больших чисел. Следует подчеркнуть, что все перечисленные стадии длились десятки и сотни миллионов лет, в течение которых сменяли друг друга миллиарды поколений последовательно усложнявшихся предорганизмов. Начался третий этап эволюции – этап развития многоклеточных организмов. При этом наметились три ветви развития живых организмов: - царство животных; - царство растений; - царство грибов. Ясно, что все три царства имеют свои стадии эволюции. Тем не менее, первые стадии у них если не полностью совпадают, то достаточно похожи друг на друга. Назовём три из них. Первая стадия – стадия фагоцителл [55]. Первоначальной топологической конструкцией многоклеточных организмов была труба (цилиндр, фагоцителла). Такая конструкция позволяла лучше защищаться от внешних воздействий и усовершенствовать условия метаболизма. Действительно, простейшая фагоцителла имела два слоя – наружные клетки специализировались на защите организма, а внутренние – на переваривании пищи. Следующая стадия – разделение времени жизни организма на две фазы – зародышевую и взрослую (репродуктивную). Зародышевый организм был в большей степени освобождён от поиска продуктов питания в окружающей среде – этими продуктами его обеспечивали родители. Поэтому, например, у предков животных зародыши представляли собой топологическую фигуру типа стакана или мешка – цилиндр с одним торцевым отверстием – бластопором. И, наконец, третья общая стадия – появление межклеточного механизма управления (регуляции, обратной связи). Первоначально это была трофическая система управления – управляющие клетки обеспечивали усиленное питание необходимыми продуктами тех участков организма, которым предстояло передвигаться или как – либо ещё взаимодействовать с окружающей средой. Затем появился гормональный способ – управляющие клетки инициировали те или иные процессы в управляемых путём направления особых ферментов – гормонов. И, наконец, у предков животных – электрофизиологический способ: с помощью нервной системы. У растений и грибов в дальнейшем развивались только первые два способа, а у животных – все три, причём наиболее интенсивно – третий. Между всеми тремя царствами живых существ установилась тесная взаимосвязь. Растения являются главным поставщиком химических веществ, необходимых для жизни – аминокислот, нуклеотидов, их полимеров, липидов, сахаридов и кислорода. Грибы служат определенными ассенизаторами, преобразуя погибающих или погибших организмов в питательные вещества жизни других организмов. И, наконец, животное царство является вершиной развития природы, включающей в себя наиболее сложные, наиболее емкие с точки зрения информации объекты. Следует отметить, что, несмотря на то, что многоклеточные более приспособлены к сохранению жизни, чем одноклеточные, последние путем длительной эволюции приспособились и к современной жизни. В настоящее время известно более 10 тысяч видов простейших [38]. Некоторые из них достигают размеров в 1 и 2 мм, например, амеба Pelomyxa palustris. Многие из них покрыты известковым панцирем (раковиной), например, радиолярии. Как видим, при развитии животного царства соблюдается основной закон накопления гармонии в диссипативных системах (см.§3.5) – закон древовидного развития. Жесткая борьба за существование способствовала не только формированию особо сложных организмов, но и дальнейшему совершенствованию менее развитых и их связи с более развитыми особями. В результате возникла многоструктурная экологическая сверхсистема, включающая в себя все дошедшие до нашего времени виды живых существ. В эту систему входят не только живые организмы, образующие флору и фауну, но и окружающие их неживые структуры – гидросфера, атмосфера, суша, недра Земли, различные космические поля, окружающие Землю, особенно если они носят периодический характер. На рисунке 5.11 дано родословное дерево животного мира [53]. Как видим, развитие животного мира берётсвоё начало с фагоцителл – колоний клеток, имеющих двухслойную цилиндрическую конструкцию (наружные клетки со жгутиками исполняли функцию движения и защиты, а внутренние - пищеварительную). После этого появились сразу пять типов - "мшанки", “планкозоиды”, “кишечнополостные” и "плоские” черви. Только одна из этих ветвей дала дорогу для дальнейшего развития: плоские черви. От них появилось четыре типа. Далее наибольшее число типов возникло у ветви, берущей свое начало от кольчатых червей. Наиболее сложные, наиболее развитые виды живых организмов появились на пути развития, берущим свое начало от полухордовых. Именно эту ветвь и следует считать главным стволом развития.
Рисунок 5.11.Родословное древо животного мира
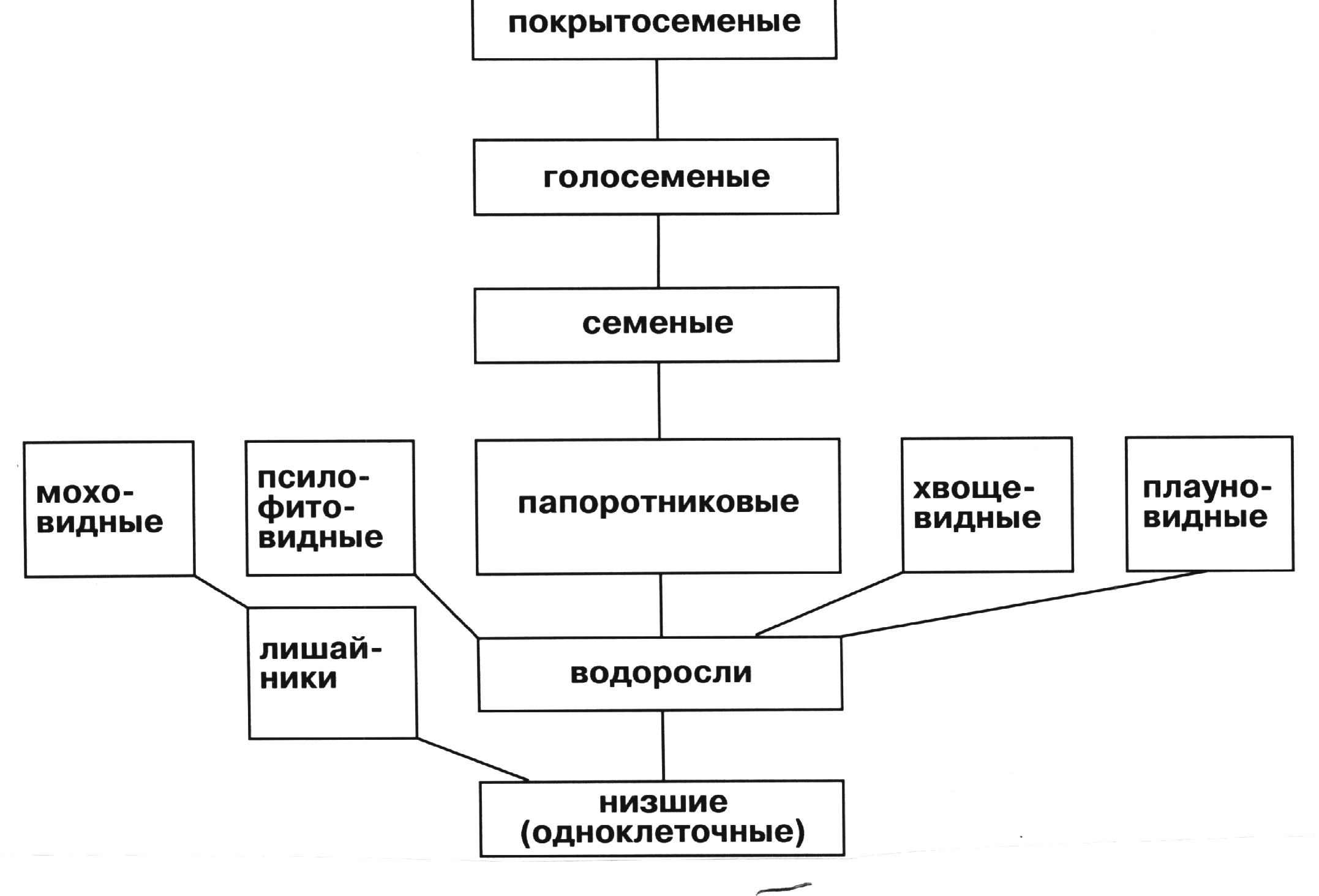
Интересно, что оба ствола развития животного царства определяются переходом от конструкции типа стакана – цилиндра с закрытым на начальной стадии жизни организма одним из торцов – к взрослому, при котором этот торец открывается, образуя либо входное, либо выходное отверстие. В первом случае появились форониды (вторичноротые), во втором – моллюски и членистоногие (первичноротые) [79]. В районе открытого торца развивался центр управления организмом. Таким образом, первый ствол развития привел к появлению в конечном счёте двухуровневого управления – спинного и головного мозга, а второй – только головного. Вместе с тем у вторичноротых возникло два узла движения: у анального отверстия (ротового в зародышевый период развития организма) и у орального. У первичноротых узел захвата пищи, на базе которого появился орган движения, только один – возле ротового(орального)отверстия. Согласно [42] развитие животного мира можно разбить на следующие периоды: -докембрийский (670-590 млн. лет тому назад) – возникновение и расцвет кишечнополостных и кольчатых червей; - кембрийский и силурийский (590-438 млн. лет тому назад) – морские беспозвоночные с твёрдой раковиной; -девонский (438-408 млн. лет тому назад) – панцирные рыбы; - карбонский (408-300 млн. лет тому назад) – хрящевые рыбы; - триасский (300-248 млн. лет тому назад) – амфибии; - пермский период (286-144 млн. лет тому назад) – пресмыкающие (рептилии); - меловой период (144- 65 млн.лет тому назад) –по-прежнему,рептилии; - кайнозойский (65 млн. лет тому назад) – млекопитающие и птицы. Параллельно развивались следующие классы животных (побочные ветви развития): - от позднего карбонского до кайнозойского (290-65 млн. лет т.н.) – насекомые; - от триасского до кайнозойского (248-2 млн. лет т.н.) – костные рыбы. Одновременно усложнялся не только сам организм, но и его "принцип действия". Химические процессы, сопровождавшие обмен веществ, передачу наследственной информации, рост и развитие организма, взаимодействие с окружающей средой, стали дополняться функциями периодического отдыха (сна), мимикрии, накопления запасов, создания жилья, коллективного труда, обмена информацией. Так, наряду с физиологией стала нарождаться психология – еще более высокая форма жизни. В ДНК половых клеток стали записываться не только гены строения органов будущего организма, но и его "принципа действия" – физиологии и психологии. Записанные в наследственной информации элементы поведения органов живых существ и их самих именуется в биологии безусловными рефлексами или инстинктами. Управление многоклеточными, особенно высокоорганизованными организмами основывается не только на наследственной информации. Память этих организмов устроена так, что она гибко перестраивает стиль поведения в зависимости от реальной среды, в которую попадает данная особь. Так возникают условные рефлексы. На клеточном уровне формирование условных рефлексов характеризуется перестройкой межнейронных связей в головном мозге [33]. Суть этого процесса заключается в появлении новых контактов “коллатерали одних нейронов - синапсы других нейронов”. В том случае, когда эти рефлексы повторяются на протяжении многих поколений по линии и мужских, и женских особей, они закрепляются в виде инстинкта. Так, например, возникли инстинкты многих домашних животных, которые без человека жить не могут, зато развили в себе именно те качества, которые нужны человеку. Надо сказать, что надолго закрепляются те инстинкты, которые несут в себе дополнительную степень гармонии (отрицательной энтропии). Дело в том, что уменьшение гармонии сразу ставит данный организм в невыгодные условия в постоянной борьбе между сохранением жизни и накоплением энтропии, о которой говорилось выше. На рисунке 5.12 дано эволюционное древо растительного царства [29]. Начало растительному царству положили одноклеточные, содержащие хлорофилл – пигмент, осуществляющий фотосинтез и благодаря ему перерабатывающий неорганические вещества в аминокислоты и нуклеотиды. Ясно, что существовали эти одноклеточные организмы в неглубоких водных участках, куда проникали солнечные лучи. Дальнейшее развитие растительного мира связано с появлением суши. Археологические исследования показывают, что растительное царство появилось на Земле всего 450 миллионов лет назад, т.е. спустя 3 миллиарда лет после появления первых живых организмов. На сушу растения вышли в конце силурийского периода – 450 - 430 миллионов лет назад. Тогда из водорослей появилось целых пять отделов – споровые, псилофитовые, плауновидные, хвощевидные и папоротниковые. При этом размножение растений от спорового вида переходит постепенно, так же, как и животных, к половому способу. Это позволяет сразу увеличить продуктивность "конструкторской" деятельности природы. Правда, для обмена половыми клетками использовался капельно-жидкостный
Родословное древо растительного царства
способ. Появление высших растений связано с возникновением "сухого" метода распространения половых клеток, среди которых особую роль играют живые организмы, в частности, насекомые [28]. Так начали зарождаться биогеоценозные сообщества. Животные питались в основном органическими веществами и кислородом, вырабатываемыми растениями, а те, в свою очередь, использовали подвижные живые организмы для своего ускоренного распространения и структурного усовершенствования. На рисунке 5.13 дано структурное древо царства грибов. Грибы играют важнейшую роль в биогеоценозе – системе, включающей в себя все существующие в настоящее время организмы. Их роль в первую очередь ассенизаторская – они ускоряют процесс перевода полураспавшихся организмов в питательную среду для вновь нарождающихся живых объектов, как растительных,так и животных [43].Общее, что объединяет все виды живых организмов, это постоянное стремление каждого из них сохранить накопленную в нем степень порядка, гармонии. Причем если животные используют для этого три способа – движение, строение организма и размножение, то представители других царств – только два последних.
Рисунок 5.13 Родословное древо царства грибов
Биосфера. Биологические организмы, возникнув примерно 3,5 ÷ 4 млрд. лет тому назад, существенно видоизменили наружный слой Земли. Суммируясь благодаря постоянному размножению, цели сохранения порядка каждого организма сформировали новую цель всего биологического мира – упорядочение всей природы этого слоя. Главная причина смены одной археологической эры другой заключена в развитии органического мира. Благодаря живым организмам изменился химический состав атмосферы. Его современное состояние – результат жизнедеятельности в основном растений. Живые организмы оказались создателями почвы и кремнеземных слоёв наружной поверхности Земли. Создавая наружную оболочку Земли, биологические системы одновременно приспосабливались к ней. В результате возникла очень сложная система взаимозависимости живых организмов (биоты) и неживой природы – биосфера. Конечно, живой мир Земли обладает весьма высокой степенью устойчивости и необходимо очень существенное изменение условий на поверхности Земли, чтобы его уничтожить. Однако даже не очень большие изменения в неживой природе вызывают катастрофическое изменение биоты. Как правило, при этом погибают менее сложные, менее развитые организмы, и за этим следует расцвет более совершенных живых существ. Так произошло в конце мелового периода, когда на смену царству рептилий пришло царство теплокровных – млекопитающих и птиц [42]. Тем не менее, главной движущей силой развития живой природы является внутренняя потребность экосистемы к самосовершенствованию, обусловленная неограниченным размножением каждой популяции, вызванным ею естественным отбором, о чём уже говорилось выше. В последние годы, особенно во второй половине 20 века в развитие биосферы вторгается новый фактор – деятельность человека и создание им внутри биосферы ноосферы – интеллектуальной оболочки Земли. Об информационных процессах, связанных с возникновением ноосферы, будет сказано ниже. Здесь остаётся подчеркнуть, что ноосфера является частью биосферы, сложившейся в конце четвертичного периода. Поэтому разрушение или даже повреждение этой биосферы несомненно скажется разрушительно на первой.
Поиск по сайту: |
 К. Во вторую – наличие гидро- и атмосферы, необходимых для жизни. В-третьих, срединное положение Земли, спасающее ее в значительной степени от "бомбардировки" окружающими Солнечную систему космическими объектами и поэтому обеспечивающее длительное бескризисное развитие.
К. Во вторую – наличие гидро- и атмосферы, необходимых для жизни. В-третьих, срединное положение Земли, спасающее ее в значительной степени от "бомбардировки" окружающими Солнечную систему космическими объектами и поэтому обеспечивающее длительное бескризисное развитие.