|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Локализация временных связей
Сначала Павлов предполагал, что временные связи образуются между корой больших полушарий, куда поступает информация о раздражении органов чувств различными сигналами, и центрами безусловных рефлексов (пищевого, Оборонительного), которые представлены на разных уровнях головного мозга, и так называемых подкорковых структурах. Например, процесс слюноотделения обеспечивается продолговатым мозгом, но пищедобывательная деятельность в целом регулируется центрами голода и насыщения в промежуточном мозге. Однако, предположение И.П. Павлова о том, что временные связи образуются между корой и подкорковыми структурами, не подтвердилось дальнейшими исследованиями.
Ученик И.П. Павлова Э.А. Асратян подвергал животных операции декортикации, то есть удалению коры больших полушарий, и исследовал у таких животных безусловные пищевые, двигательные, вегетативные рефлексы. Оказалось, что удаление коры больших полушарий сопровождается существенным снижением качества безусловных рефлексов. Это проявляется в том, что исчезают, например, различия между качеством и количеством слюны, выделяемой слюнными железами на разные виды пищи - сухари, молоко, мясо.
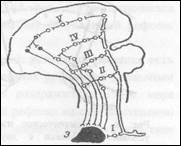
Нарушается координация движений, усиливаются реакции сердечно сосудистой системы на адреналин. Результаты этих наблюдений позволили Асратяну сделать вывод о том, что каждый безусловный рефлекс имеет, помимо подкорковых центров представительство в коре больших полушарий. Это так называемое корковое представительство безусловного рефлекса повышает качество безусловных рефлексов, осуществляемых подкорковыми центрами (продолговатым средним, промежуточным мозгом). Таким образом, центральная часть дуги любого безусловного рефлекса выглядит многоэтажной, поскольку информации от всех видов рецепторов, идущая по чувствительным (афферентным центростремительным) нервам, подвергается частичной обработке в разных отделах головного мозга. Высший же анализ этой информации осуществляется в коре больших полушарий - в корковом представительстве безусловной рефлекса (рис.7)
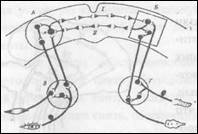
На основании данной концепции была предложена новая схема образовании условного рефлекса, представленная на рис. 8. Любой раздражитель, который мы хотим сделать условным сигналом, вызывает у животного, безусловным ориентировочный рефлекс, названный И.П. Павловым рефлексом «что такое?» Биологическое значение этого рефлекса очевидно, поскольку животного
ориентировочный рефлекс на вспышку лампочки проявится в форме двигательных реакций - повороты головы в сторону света, фиксировании лампочка взглядом. Являясь безусловным, данный рефлекс, в соответствии с концепцией Асратяна обеспечивающую быструю реали зацию этих движений, и корковое представительство, делающее эту реакцию более точной и совершенной. Известно, что подкорковая ветвь ориентировочных безусловных рефлексов на свет и звук включает средний мозг (соответственно, передние и задние холмы). Однако, информация о раздражении рецепторов сетчатки поступает I и кору больших полушарий - в корковое представительство данного ориентировочного рефлекса. Корковый зрительный центр, обработав пришедшую информацию, посылает сигналы в подкорковые центры, улучшая количество безусловного рефлекса (рис. 7). Подобные рассуждения справедливы и относительно безусловного подкрепления. Раздражение пищей рецепторов ротовой полости трансформируется в нервное возбуждение, передаваемое по лицевому, языкоглоточному и другим нервам в пищевые центры продолговатого мозга, что вызывает выделение слюны. Вместе с тем, информация о раздражении рецепторов передаётся и в корковое представительство пищевого безусловного рефлекса, что, как уже указывалось, делает реакцию подкорковых центров более точной и совершенной. Таким образом, если сочетать два сигнала – свет пищевое подкрепление, то временная связь образуется, согласно представлениям Асратяна, между корковыми представительствами двух безусловных рефлексов: ориентировочного рефлекса на свет и пищевого рефлекса на раздражение пищей рецепторов ротовой полости. В силу этого свет становится сигналом кормления, то есть всякий раз, когда включается лампочка, свет вызывает не только свойственный ему безусловный рефлекс "что такое?", но и реакцию слюноотделения. Таким образом, по современным представлениям, условный рефлекс есть корковый синтез двух или нескольких безусловных рефлексов(условным сигналом может быть не один, а несколько раздражителей). По мере укрепления условного рефлекса ориентировочный рефлекс на свет постепенно затухает и в полной мере проявляется сигнальное значение света, то есть свет тает слюноотделение. При выработке условных рефлексов выделяют две стадии – стадию образования и стадию упрочения временных связей, то есть закрепления условного рефлекса. В психологическом плане этим стадиям соответствуют понятия кратковременной и долговременной памяти. Впервые в конце 19 века русский психиатр С.С. Корсаков наблюдал синдром разъединения двух видов
памяти при алкогольной интоксикации, приводящей к белой горячке. Оказалось, что выйдя из состояния белой горячки, человек не помнил непосредственных событий, предшествовавших интоксикации, но помнил события, далеко отстоящие от приступа болезни. Эти наблюдения позволили предположить, что механизмы кратковременной и долговременной памяти различны, поскольку шоковое состояние человека сопровождается потерей кратковременной, но не долговременной памяти. Это явление, получившее название «синдрома Корсакова», наблюдается при шоковых состояниях, вызываемых различными воздействиями (алкогольная и другая интоксикация, черепно-мозговые травмы, связанные с потерей сознания и коматозным состоянием и др.). Подобный синдром можно наблюдать и в опытах по выработке условных рефлексов у лабораторных животных. Крысам в разные сроки после сочетании условного и безусловного сигналов наносили электрические раздражения, вызывающие судорожные припадки. Оказалось, что если электрошок вызывался через 20 сек - 60 мин после выработки условного рефлекса. то животное, выйдя из состояния шока, «не помнило» значения условного сигнала. Если же до нанесения раздражения током проходило более одного часа, то, выйдя из состояния шока, крыса правильно реагировала на условный раздражитель. Таким образом, эти опыты наглядно показали, что процесс образования условного рефлекса состоит из двух фаз. Первая - образование временных связей, обеспечивающее кратковременную память. Вторая упрочение (консолидация) временных связей, обеспечивающее долговременную память. В данном конкретном случае для перехода кратковременной памяти в долговременную необходимо не менее одного часа. Каковы механизмы кратковременной и долговременной памяти? Как уже указывалось, при образовании условного рефлекса наблюдается встречная иррадиация возбуждения между двумя центрами, что приводит к «проторению пути» (баиунг-доминанта).Разберем нейрофизиологический механизм этот явления, составляющего физиологическую основу кратковременной памяти. Между двумя центрами, например, зрительным и пищевым, расположено огромное количество нейронов с многочисленными нервными отростками которые до выработки условного рефлекса не в состоянии проводим возбуждение. Образование условного рефлекса состоит в том, что «потенциалъные» синапсы, лежащие между двумя центрами, превращаются
в «актуальные». Иначе говоря, неработающие синапсы в процессе неоднократных сочетаний условного и безусловного сигналов становятся работающими, то есть способными к проведению возбуждения. Процесс актуализации синапсов осуществляется несколькими путями. Во-первых, происходит переориентация нервных отростков, в результате чего между нервными центрами образуется как бы дорожка, состоящая из системы упорядоченных нейронов и синапсов. Но наличие такой дорожки не является гарантией проведения нервного импульса, поскольку расстояние между пресинаптической и постсинаптической мембранами может быть слишком большим. В результате этого медиатор, выделяемый пресинаптической мембраной, проходя через слишком широкую синаптическую щель, теряется пространстве. Устранение этого недостатка происходит за счет утолщения (набухания) пре- и постсинаптических мембран. Набухание мембран, уменьшая ширину синаптической щели, приводит к тому, что медиатор достигает постсинаптической мембраны и вызывает ее деполяризацию и возникновение потенциала действия, то есть синапс начинает проводить возбуждение. Еще один из путей актуализации синапсов связан с образованием миелиновой оболочки в случае, если часть нервных волокон в системе нейронов, образовавших «дорожку» между двумя центрами, являются безмякотными. Как известно, возбуждение по безмякотным волокнам проводится медленно и с потерей энергии нервного сигнала (декрементный тип проведения возбуждения). В силу этого к пресинаптической щели подходит небольшой по амплитуде потенциал действия, который стимулирует выделение синаптическую щель небольшого количества медиатора, неспособного возбудить постсинаптическую мембрану. В результате возбуждение через синапс не передается. При выработке условного рефлекса происходит образование миелиновых оболочек вокруг безмякотных нервных волокон помощью клеток опорной ткани (глии). Сформированное мякотное волокно характеризуется быстрым и бездекрементным проведением возбуждения. Теперь потенциал действия, не теряя своей амплитуды, подходит к пресинаптической мембране, через которую выделяется количество медиатора, достаточное для возбуждения постсинаптической мембраны. Таким образом, сигнал передается с одного нейрона на другой. Следовательно, в основе формирования кратковременной памяти лежат физиологические процессы актуализации синапсов на стадии образования условных рефлексов.
Долговременную память,в основе которой лежит закрепление условного рефлекса, связывают с химическими механизмами. Мак Коннел провел опыты на плоских червях планариях. У планарий вырабатывали пищевой условный рефлекс на свет, после чего обученную планарию разрезали пополам. Из каждой половины вследствие регенерации образовывались взрослые особи, у которых подобные условные рефлексы вырабатывались значительно быстрее. Опыты модифицировали, приготавливая из «обученных» планарий суспензию и вводя ее необученным планариям. Результат оказался такой же - у планарий -реципиентов ускорялась выработка условных пищевых рефлексов на свет. Такие результаты получались и при скармливании обученных планарий необученным. Возникло предположение, что в основе упрочения временных связей лежат химические изменения в молекулах ядерной РНК. Действительно, если одну из половинок перерезанной обученной планарий поместить в морскую воду, куда добавлен фермент РНК-аза, расщепляющий РНК, то выросшая в таком растворе планария не проявляет способности к ускоренной выработке условного рефлекса на свет. В результате подобных экспериментов было развито представление о том, что информация, поступающая в нервные клетки коры больших полушарий, приводит к изменениям структуры молекулы ядерной РНК, вызывая изменения последовательности в соединении нуклеотидов - аденина, гуанина, цитозина и урацила. Автором химической теории долговременной памяти является Хиден. Возможность участия РНК в процессах запоминания подтверждается «большой информационной емкостью» РНК. Если использовать все перестановки и комбинации нуклеотидных элементов в цепочках РНК молекул нервной клетки, то получится число 1015 - 1020, что сопоставимо с объемом человеческой памяти, выражаемой в битах (единицах) информации. Эксперименты, подобные опытам Мак Коннела на планариях, были проведены на других видах животных. Результаты показали, что введение суспензии мозга обученных животных необученным повышало у последних скорость образования соответствующих условных рефлексов. Оказалось, что у пожилых людей с ослабленной памятью введение раствора чистого РНК улучшало процессы запоминания. Эти данные свидетельствуют в пользу химического механизма долговременной памяти, связанного с изменениями структуры макромолекул РНК. Следует отметить, однако, что процессы, обеспечивающие физиологическую основу кратковременной и долговременной памяти, еще не расшифрованы до конца. Несомненно, что память обеспечивается комплексными, системными механизмами, среди которых немаловажную роль играют биофизические, химические процессы, различные медиаторы и гормоны.
Поиск по сайту: |