|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Протонефридии и метанефридии
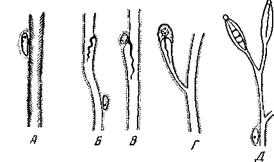
Протонефридии впервые появляются у турбеллярий; первоначально это органы осморегуляции, которые, однако, очень скоро вступают в связь с экскреторными клетками и приобщаются к выделительной функции. У кишечнополостных нефридиев нет и гомологи их неизвестны. Точно так же их нет и у Acoela и Xenoturbellida. У Polyclada они встречаются редко, развиты слабо и мало изучены. Во всех остальных отрядах турбеллярий они большей частью имеются. Анатомически наиболее примитивно построены протонефридии большинства триклад. Они состоят из восьми тонких продольных каналов, залегающих в паренхиме. Четыре лежат на спинной стороне, четыре – на брюшной (рис. 140, А).Все они соединены между собой поперечными комиссурами и дают многочисленные ветви, распадающиеся на сеть капилляров, пронизывающих все тело. От главных каналов по направлению к коже отходят многочисленные короткие выводные протоки, открывающиеся наружу маленькими порами. Экскреторной функции протонефридии триклад не несут. У вышестоящих или более мелких форм мы наблюдаем архитектоническое упрощение, связанное с олигомеризацией: уменьшается и число продольных каналов, и число поперечных комиссур, и число наружных отверстий (рис. 140). Это явление имеет место уже у карликовой триклады Pentacoelum caspicum. У Alloeocoela Holocoela имеются уже не восемь, а четыре канала, всего две поперечные комиссуры и три-четыре пары наружных отверстий. У Rhabdocoela остаются всего два канала с двумя, а иногда и с одним наружным отверстием; поперечных комиссур совсем нет. У Notandropora (сем. Catenulidae) сохраняется всего один продольный канал. Каналы сложно ветвятся и дают сеть капилляров. Трематоды по архитектонике своих протонефридиев стоят на одном уровне с Rhabdocoela, т.е. имеют один или два канала без поперечных комиссур, с одним или двумя наружными отверстиями. У цестод встречается большое количество отверстий и поперечные комиссуры: между стволами, так что многие из них по уровню организации своих цротонефридиев приближаются к трикладам. У TyphloplanicJse (Bhabdocoela) наружное отверстие протонефридиев сочетается с ротовым отверстием (Mesostomatini, Typhloplaninae) или с половым (Rhynchomesostomam). В гистологическом отношении (рис. 141) наиболее простую структуру нефридиев описывает Е. Reisinger для Protomonotresis centrophora У Baikalarctia (Holocoela) строение канала то же, но реснички располагаются не сплошь, а лишь тремя продольными рядами. Дальнейшую ступень в редукции и дифференциации ресничного аппарата протонефридиев мы встречаем у некоторых Rhabdocoela (рис. 141, В, В);стенки канала построены здесь по-прежнему, но ресничек больше нет. Лишь кое-где на утолщениях стенки имеются длинные пучки слепленных вместе ресниц, которые лежат в просвете канала, и, извиваясь, гонят вдоль него воду. Если мы представим, что подобный пучок ресниц углубится в выпячивание стенки канала, мы получим типичный концевой аппарат, свойственный большинству протонефридиев.
Рис. 140. Схемы анатомического строения протонефридиального аппарата платод А – Euplanaria polychroa (Triclada Paludicola), поперечный разрез: 1, 2, 3 и 5– главные выделительные каналы – дорсолатеральный, дорсомедиальный, вентролатеральный и вентро-медиальный; 4 – брюшные нервные стволы (по Миколецкому); Б – Baihalarciia gulo
Рис. 141. Вододвижущие приспособления в нефридиях турбеллярий А – равномерно покрытый ресничками выделительный канал Protomonotresis Получается колбочка, на дне которой сидит «мерцательное пламя», т.е. пучок слипшихся ресничек, торчащий в полость канала и находящийся в непрерывном движении (рис. 141, Г, Д).Для Digenea характерно ограниченное число и правильное расположение концевых аппаратов нефридиев. Такова высшая ступень дифференцировки протонефридиев у плоских червей. Какова же функция этого аппарата? Обычно ему приписывают выделительную функцию. Однако первоначальной и основной функцией протонефридиев является не экскреция, а регуляция осмотического давления в теле животного, и прежде всего удаление воды, непрерывно насасываемой из окружающего гипотонического раствора, каковым, например, является для всех животных пресная вода. Поэтому протонефридии очень слабо развиты у морских турбеллярий по сравнению с пресноводными. У триклад протонефридии еще не имеют к выделительной функции никакого отношения. У части Rhabdocoela атроциты располагаются по ходу главных каналов протоиефридиев и отдают скопляющиеся в них экскреты току жидкости, непрерывно протекающей по каналам. Таким образом, экскреторные элементы вступают в связь с протонефридиями. Наконец у некоторых Rhabdocoela (Gyratrix, Acrorhynchus) и у Stenostomum эпителий самих каналов в известных его участках приобретает самостоятельную экскреторную функцию. Только с этого момента протонефридии становятся экскреторными органами. Таким образом, у высших турбеллярий они несут двойную функцию: не только осморегуляторную, но и выделительную. Концевые аппараты экскреторной функции не несут. Интересен вопрос о первоначальной, наиболее примитивной форме протонефридиев. Протонефридии всегда образуются за счет эктодермы. Строение их у Protomonotresis свидетельствует, что первоначально они представляют впячивание наружного мерцательного эпителия. Поэтому первоначальным типом протонефридиев естественно было бы считать множество отдельных трубочек, открывающихся наружу самостоятельными отверстиями, наподобие трахейной системы онихофор. Однако нефридиальная система такого типа среди плоских червей нам неизвестна. Только у аннелид имеется много пар метамерно расположенных, независимых, слабо ветвящихся нефридиев. Нефридии триклад представляют следующий этап. Первоначально независимые кустики протонефридиев слились уже в единую систему, сохранив, однако, множественность наружных отверстий. Все остальные типы нефридиального аппарата плоских червей представляют дальнейшие стадии интеграции и архитектонического упрощения этого первоначального типа. При этом бросается в глаза некоторый формальный параллелизм с интеграцией и архитектоническим упрощением трахейной системы Tracheata. Этот параллелизм, очевидно, обусловлен лишь тем, что в обоих случаях мы имеем систему тонких трубочек, представляющих впячивание покровов и функция которых заключается в установлении притока или оттока веществ между наружной средой и тканями тела. За пределами плоских червей протонефридии довольно широко распространены. Из ближайших родственников плоских червей они встречаются в обоих отрядах Gastrotricha. Затем мы их находим у коловраток (рис. 142, А),киноринх, у части скребней (рис. 142, Б),а также у приапулид (рис. 142, В).Снабжены протонефридиямии немертины. Таким образом, протонефридии свойственны всем группам сколецид, за исключением нематод и волосатиков, вообще лишенных ресничек. Отметим, что у нематод осморегулирующую функцию взяли на себя одноклеточные кожные железы, а именно – так называемые шейные железы; отчасти они несут и экскреторную функцию; последнюю, как и у турбеллярий, несет также и кишечник. У Ascaris шейные железы тянутся по боковым линиям почти вдоль всего тела и открываются наружу общим протоком на вентральной стороне близ переднего конца тела. Парные протонефридии Gastrotricha Chaetonotoidea и киноринх в связи с крошечными размерами этих животных имеют каждый всего по одной концевой колбочке. В то время как протонефридии плоских червей представляют сложную систему разветвлений, пронизывающую все тело, протонефридии коловраток и немертин представляют собой короткие и толстые каналы с немногими короткими разветвлениями, покрытыми большим количеством тесно сидящих концевых аппаратов. У крупных коловраток (Asplanchna, рис. 142, А)число этих колбочек достигает иногда нескольких десятков, у мелких их бывает по 2–8 на каждый протонефридий. Отличие между протонефридиями плоских червей и таковыми коловраток и немертин объясняется тем, что протонефридии плоских червей погружены в паренхиму, внутри которой диффузия воды и экскретов трудна, и потому дренирующая система должна проникать во все концы тела. Наоборот, у коловраток, нефридии погружены в обширный схизоцель, омывающий все органы, и заполняющая его жидкость служит посредствующим звеном между тканями, с одной стороны, и протонефридиями – с другой. Точно так же и у немертин протонефридпи погружены в сосуды кровеносного аппарата, который сам дренирует все тело, чем создается возможность концентрации нефридиев. Таким образом, концентрированная форма протонефридиев коловраток и многих немертин, в противоположность охватывающим все или почти все тело протонефридиям плоских червей, связана с более совершенным распределительным аппаратом двух вышестоящих групп. У большинства коловраток выделительное отверстие сочетается с половым и заднепроходным, у скребней и приапулид протонефридии впадают в половые протоки. Подобное сочетание выделительного отверстия с половым мы уже видели у Rhynchomesostomum среди прямокишечных турбеллярий. Пара типичных протонефридиев нередко встречается у трохофорных личинок аннелид и моллюсков (рис. 143, А, Г),причем у моллюсков они только на стадии личинки и встречаются. Взрослые моллюски протонефридиев никогда не имеют. У аннелид нефридии трохофоры при метаморфозе большей частью также редуцируются, но зато при дальнейшем развитии в каждом сегменте тела образуется по паре нефридиев.
Рис. 142. Протонефридии сколецид А – Asptanchna (Rotatoria): 1 – подвесочный тяж; 2– жгуты, направленные в полость тела; Б– Hamaniella microcephala (Acanthocephala): 1– ядра стенок нефридия; 2 – канал нефридия; 3 – отдельные концевые аппараты с мерцательным пламенем; Таким образом, в прототипе аннелидам свойственно весьма характерное расположение нефридиальной системы: множество метамерных пар нефридиев, погруженных в целомические мешки, по одному нефридию на каждый мешок. В простейшем случае каждый из этих нефридиев построен примерно по типу нефридиев коловраток или немертин, т.е. представляет кустик каналов, торчащих в полость тела и покрытых концевыми аппаратами. Что касается концевых аппаратов, то у взрослых полихет они претерпели дальнейшее изменение и представляют трубчатые клетки, заканчивающиеся ядросодержащим утолщением и содержащие, вместо мерцательного пламени, единственный длинный жгут (рис. 143, Б).За свою трубковидную форму клетки эти названы соленоцитами. Наиболее просто устроенными протонефридиями обладают представители сем. Phyllodocidae, например виды рода Phyllodoce (рис. 143, В).Из других взрослых полихет типичные, замкнутые протонефридии, снабженные соленоцитами, встречаются у Nephthydidae и Glyceridae. Более существенное видоизменение нефридиев представляет та их форма, которую Э. Гудрич предлагает обозначать как метанефридии. Метанефридии встречаются у взрослых особей Nereidae, Hesionidae и Polygordius. Личинка Polygordius имеет две пары типичных протонефридиев с концевыми аппаратами. Первая пара погибает вместе со всеми провизорными органами личинки. Вторая пара теряет свои концевые аппараты, но дистальная часть ее входит в состав первой пары нефридиев взрослого червя. При образовании сегментов тела в каждом из них образуется по паре нефридиев, состоящих из небольшого извитого канальца, лежащего в целоме и открывающегося одним концом наружу, другим концом, при помощи маленького нефростома – в полость предшествующего сегмента. Отсутствуют нефростомы лишь у одной пары дефинитивных нефридиев. Благодаря наличию нефростома нефридии Polygordius и других сходных в этом отношении форм (рис. 143, В)очень напоминают целомодукты, с которыми их до исследований Гудрича и смешивали. Анатомическое отличие между теми и другими выражается у полихет главным образом в том, что нефростом представляет узкое отверстие, тогда как целомодукты обычно открываются в целом широкой мерцательной воронкой. Эмбриологические различия более определенны: нефридии образуются за счет эктодермы (наружной кожи) или же за счет эктомезодермальных нефробластов и растут от периферии в сторону целома. Целомодукты образуются за счет целомического эпителия и растут по направлению к наружной коже. Эктодермальное происхождение нефридиев, в частности, доказано для Polygordius. У олигохет они образуются за счет эктодермальных телобластов, как доказано для Criodrilus. Из пиявок то же самое доказано для Nephelis. До работ Goodrich выделительные органы беспозвоночных разделяли на протонефридии, снабженные концевыми колбочками, и метанефридии, снабженные воронками, открытыми в целом. Ныне такое разделение совершенно неприемлемо, так как оказалось, что каналы с воронкой, открывающиеся в целом, по природе своей бывают различны: одни из них являются нефридиями, другие – целомодуктами. Несмотря на то, что Goodrich уже более полвека назад распутал эти взаимоотношения и данные его были подтверждены рядом авторов, до сих пор в популярной литературе, а иногда и в учебниках повторяется старое смешение понятий. В настоящее время термины «протонефридии» и «метанефридии» следует применять для двух типов истинных (эктодермальных) нефридиев, обозначая замкнутые нефридии типа Phyllodoce как протонефридии, а снабженные нефростомами типа Polygordius – как метанефридии. Настоящие эктодермальные нефридии в той или иной форме встречаются у всех полихет. Однако они большей частью вступают в ту или иную связь с целомодуктами и совершенно от них независимы только в сем. Capitellidae, представители которого имеют по одной паре метанефридиев и по одной паре целомодуктов в каждом сегменте. В других семействах независимые нефридии бывают либо при редукции целомодуктов (Nereidae, Glyceridae и другие), либо у неполовозрелых особей с еще неразвитыми целомодуктами, либо в отдельных сегментах тела, в которых целомодукты отсутствуют, например, в передних бесплодных сегментах Poecilochaetus (сем. Disomidae отряда Spiomorpha).
Рис. 143. Нефридии трохофорных личинок и полихет А – трохофора Polygordius: 1– ядро концевой клетки; 2– стенка канала нефридия; 3 – жгуты 4 – протоки в выростах; 5– концевой клетки (по Goodrich); Б – Phyllodoce parelli, конец одной изветвей протонефридия с соленоцитами (по Goodrich); В– Trypanosyllis, проксимальный конец открытого нефридия: 1– диссепимент; 2 – нефростом (по Goodrich); Г– зародыш Limnea (Gastropoda): 1– концевая клетка; 2 – наружное отверстие; 3– эктодерма (по Meisenheimer) Олигохеты имеют обычно по одной паре нефридиев, снабженных нефростомами (рис. 144, А),в каждом из сегментов тела, кроме ларвальных. Но у некоторых крупных наземных форм первоначальные парные зачатки нефридиев зародыша дают начало большому количеству самостоятельных микронефридиев (сем. Perichaetidae, Acanthodrilidae, Cryptodrilidae и др.). У некоторых видов происходит умножение воронок при сохранении общего протока нефридия, например, у Thamnodrilus crassus (Glossoscolecidae). У Allolobophora antipae (Lumbricidae) часть нефридиев каждой стороны сливается в продольный канал, и оба канала впадают в заднюю кишку – любопытная конвергенция с первично-почечным протоком позвоночных. Заметим, однако, что выделительный аппарат позвоночных, образованный целомодуктами, не гомологичен выделительному аппарату олигохет, образованному нефридиями. Экологический смысл этого приспособления у олигохет также совершенно иной и сводится к экономии воды, которая в кишечнике частично всасывается, а экскреты выделяются в обезвоженном виде. Аналогичное явление широко распространено среди тропических земляных червей сем. Megascolecidae. Их нефридии, вместо того, чтобы открываться наружу, впадают в кишечник, у некоторых форм только в задних сегментах тела, у других – в большей части сегментов и, наконец, у Ттаvoscolides – во всех. У более примитивных форм немногие открывающиеся в кишечник пары нефридиев открываются туда непосредственно; у Megascolex campester нефридии впадают в пару мочеточников, лежащих по бокам спинного кровеносного сосуда; в каждом сегменте они соединяются поперечным протоком и открываются в кишечник одной парой отверстий (рис. 144, В).Максимальная степень интеграции нефридиального аппарата из изученных видов наблюдается у Travoscolides: он имеет непарный мочеточник, спустившийся в тифлосоль и принимающий в себя метамерные нефридии, открываясь в кишечник непарными метамерными порами. Так же соединяются продольными протоками нефромиксии теребеллиды Lanice conchilega; три предиафрагмальных нефридия каждой стороны соединены между собой и четыре постдиафрагмальных – между собой; однако открываются они все наружу, а не в кишечник; живущей в море полихете экономия воды явно не нужна. У Octochaetus Acanthodrilus некоторых других олигохет несколько пар нефридиев впадает в глотку. Octochaetus имеет, кроме того, пучки микронефридиев, впадающие непосредственно в заднюю кишку, что повело к неоправданным попыткам рассматривать мальпигиевы сосуды членистоногих как видоизмененные нефридии. У паразитической олигохеты Branchiobdella число нефридиев низводится до двух пар, у некоторых наидид нефридии полностью отсутствуют. У ряда олигохет из различных семейств наблюдается редукция нефростома и образование замкнутых нефридиев – явление, широко распространенное у пиявок. Нефридии пиявок (рис. 144, В)обычно слепо замкнуты на проксимальном конце. У слепого конца нефридия находится мерцательная воронка, открытая в целом и ведущая в небольшую капсулу; капсула непосредственно примыкает к слепому концу нефридия. Полость капсулы содержит большое количество фагоцитов. Гомология воронки не совсем ясна: одни приписывают ей целомическое происхождение и считают ее гомологом ресничных органов целома полихет, другие считают ее гомологом нефростома. У очень многих Ichthyobdellidae (Rhynchobdellea) происходит такое же умножение нефридиев, которое мы видели среди олигохет у Perichaetidae и других, но у ихтиобделлид все эти нефридии остаются в соединении между собой, образуя густую сеть разветвлений в каждом сегменте. Их и здесь обозначают как плектонефридии. Некоторые авторы считают плектонефридии первичным типом нефридиального аппарата аннелид, приравнивая их к разветвленным нефридиям плоских червей. Однако и чисто морфологически совокупность отдельных небольших нефридиев примитивнее, чем интегрированная система каналов (см. выше, о нефридиях плоских червей), и с исторической точки зрения легче себе представить, что плектонефридии пиявок развились вторично из простых нефридиев большинства аннелид, ибо они встречаются в отдельных, высоко специализованных группах и полностью отсутствуют в примитивных классах, в частности у полихет. За пределами аннелид настоящие нефридии встречаются очень редко. Они имеются у Kamptozoa, Phoronis (рис. 145, А). Ни у взрослых моллюсков, ни у членистоногих настоящих нефридиев никогда не бывает, хотя Е.S. Goodrich (1946) и рассматривает выделительные железы ракообразных как нефридии. Точно так же нет настоящих нефридиев ни у кого из вторичноротых, кроме ланцетника. Наличие в нефридиях ланцетника типичных соленоцитов (рис. 146, В)было доказано Е.S. Goodrich, как и отсутствие у них нефростома, Е.S. Goodrich показал, что в действительности нефридии ланцетника развиваются из одноклеточных зачатков, попарно лежащих в каждом сегменте между целомическим мешочком и кишкой. Исходные нефробласты, по мнению Goodrich, образуются в зоне роста зародыша за счет эктодермального или эктомезодермального материала и потомки их достаются по одной паре каждому сегменту. При развитии нефридия соленоциты впячиваются в целом, нефропор открывается в жаберную кишку. Наличие настоящих нефридиев у ланцетника при отсутствии их у всех остальных вторичноротых – факт в высшей степени удивительный. Резкая обособленность типичных первичноротых и вторичноротых не подлежит сомнению. Как мы видели, и те и другие независимо выводятся из кишечнополостного прототипа. Мало того, независимо друг от друга из этого прототипа выводятся и разные группы первичноротых (сколециды, трохофорные животные). Ни у кого из современных кишечнополостных протонефридиев нет. У современных плоских червей мы видим все стадии развития протонефридиев – от простых, выстланных мерцательным синэпителием трубок, до органов, снабженных характерными концевыми аппаратами. В то же время ясно, что плоские черви не дали начала ни одной группе животных за пределами сколецид. Все это говорит в пользу независимого возникновения протонефридиев в различных стволах Bilateria.
Рис. 144. Нефридии Glitellata А – схема нефридия дождевого червя: 1– нефростом; 2 – извилистый канал; 3– диссепимент; 4– выделительная пора (по Meisenheimer); Б– концевой аппарат нефридия Clepsine: 1– воронка; 2 – капсула; 3– лейкоциты; 4– концевая клетка нефридия со слепо оканчивающимся разветвленным протоком (по Meisenheinier); В– схема строения выделительных органов Megascolex campester (Ollgochaeta) позади 20-го сегмента тела: 1– спинной кровеносный сосуд; 2 – боковой надкишечный выделительный проток; 3– поперечные соединения между обоими выделительными протоками; 4– воронка нефридия;
Рис. 145. Протонефридии, встречающиеся за пределами сколецид, аннелид и моллюсков А – схематический фронтальный разрез через актинотроху (личинка Phoronis): 1– теменная пластинка; 2– пищевод; 3– желудок; 4– щупальца метатроха (личиночные щупальца); 5– туловищный целом; 6– кишечник; 7 – телотрох; 8 – передний отдел целома; 9– нефридий, 10 – соленоциты; 11– схизоцель (по Goodrich); Б – часть нефридия Amphioxus lanceolatus (Acrania), полусхематически: l – ядра соленоцитов; 2– жгуты; 3– канал нефридия; 4– трубка соленоцита (по Goodrich)
Поиск по сайту: |