|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
СТРОЕНИЕ НЕРВНОЙ И ЭНДОКРИННОЙ СИСТЕМЫ АННЕЛИД И ЧЛЕНИСТОНОГИХ
Нервная и эндокринная система аннелид
У некоторых примитивных аннелид, например у Dinophilus, у Aeolo- soma, брюшные стволы широко расставлены и соединены длинными поперечными комиссурами, так что возникает «лестничная нервная система». Таким образом, в пределах всего подтипа аннелид централизация нервного аппарата делает лишь небольшие успехи по сравнению с наиболее примитивными представителями этой группы. Лишь у высших представителей аберрантного, олигомерного отряда Myzostomida вся брюшная цепочка оказывается слита в одну сплошную нервную массу. От каждого из ганглиев брюшной цепочки полихет отходят, как правило, 3 пары периферических нервов; но при значительном увеличении длины отдельных сегментов, как, например, в семействе Maldanidae (Drilomorpha), число периферических нервов в каждом невросомите увеличивается. В типичном случае передняя и задняя пары нервов – преимущественно двигательные, средняя – преимущественно чувствительная. Оба нерва каждой из двигательных пар соединяются между собой на спинной стороне, образуя таким образом кольцевые нервы. У блуждающих полихет, как указано выше, имеется при основании каждой параподии небольшой подиальный ганглий, соединяющийся с нервом средней пары, а через него – с брюшной цепочкой. В семействе Amphinomidae (Nereimorpha) все подиальные ганглии каждой стороны соединены также и непосредственно между собой при помощи продольных коннективов, известных под именем латеральных нервов
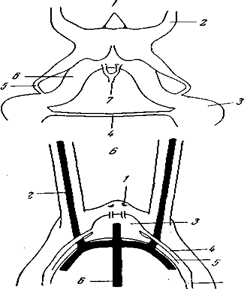
Рис. 95. Центральный нервный аппарат полихет А – Nereis virens (Polychaeta), передний конец тела (из Догеля); Б – Sabellaria alveolata (Polychaeta Hermellidae), передний конец тела (по К.E. Johansson): 1 – перистомиальные цирры; 2 – нухальные органы; 3 – окологлоточные коннективы; 4 – ганглии перистомия; 5 – брюшная нервная цепочка; 6 – нерв перистомия; 7 – мозг; 8 – глаза; 9 – пальпы; 10 – антенны; 11 – экскреторные нефромиксии в тораксе; 12 – половые нефромиксии; 13 – брюшная нервная лестница со сближенными коннективами в тораксе; I–V – пять пар нервов каждого невросомита, из которых II несет параподиальный ганглий; В – Hermodice (Polychaeta, Amphinomidae), схема нервного аппарата передней части тела: 1 – мозг; 2 – параподиальные ганглии 1-го туловищного сегмента; 3 – комиссуры I и II пары брюшных ганглиев, огибающие глотку; 4 – боковой нерв; В отличие от амфиномид и сферодорид ни у кого из других блуждающих полихет продольных коннективов между всеми подиадьными ганглиями не известно.
Рис. 96. Мозг сидячих полихет А – схема строения мозга Serpula vermicularis (вид с дорсальной стороны): 1 – передние медиальные нервы; 2 – нервы к пальпам; 3 – 1-я пара ганглиев брюшной цепочки; 4 – их комиссура; 5 – спинные корешки окологлоточных коннективов; 6 – брюшные корешки окологлоточных коннективов; 7 – нервы к спинному сосуду; Б – схема взаимоотношений мозга и сосудов головы у Stylarioides (сем. Chlorhaemidae): 1 – глаза; 2 – сосуды пальпов; 3 – мозг; 4 – спинные корешки окологлоточных коннективов; 5 – брюшные корешки окологлоточных коннективов; 6 – спинной сосуд; 7 – окологлоточные коннективы (по К.Е. Johansson)
У некоторых блуждающих полихет непосредственные соединения между подиальными ганглиями все же имеются, но лишь на протяжении нескольких первых сегментов, и отсутствуют в остальных; так, у Nereis соединены между собой подиальные ганглии 4 передних сегментов, несущих тентакулярные цирры; считает это соединение следствием цефализации. У большинства сидячих полихет подиальных ганглиев нет или встречаются их рудименты. Мозг аннелид, в примитивных случаях развивающийся из множества самостоятельных зачатков (Lopadorhynchus), у многих других полихет образуется из одного зачатка, а у олигохет – даже за счет общего зачатка с брюшными стволами. Последний способ развития несомненно является вторичным. Независимо от способов образования мозг взрослой аннелиды представляет монолитное образование, и лишь при сильном развитии самого мозга и иннервируемых им органов чувств (у блуждающих полихет) в нем обособляются три отдела: передний мозг, иннервирующий пальпы; средний мозг, иннервирующий глаза и тентакулы; задний мозг, иннервирующий нухальные органы. Ввиду того, что зачатки пальпов образуются по бокам теменной пластинки трохофоры, можно принять, что передний мозг возник за счет зачатков, расположенных вокруг аборального полюса, а задний мозг, как и обслуживаемые им нухальные органы, – в спинном (D) квадранте верхнего полушария; таким образом, задний мозг вполне оправдывает свое название. Чем сильнее развиты какие-либо органы чувств, тем сильнее развит и соответствующий отдел мозга: амфиномиды с их громадными нухаль-ными органами имеют большой задний отдел с хорошо развитыми ассоциативными центрами (гломерулами). У Alciopidae (Phyllodocemorpha), имеющих большие и относительно совершенные глаза, хорошо развит средний мозг и зрительные центры. У нереид и афродитид, обладающих хорошо развитыми пальпами, сильно развит передний мозг. Eunicidae имеют из всех полихет наиболее сложно устроенную глотку, с вентральным слепым мешком и наиболее сложным глоточным отделом нервного аппарата, и это обстоятельство тоже усиливает развитие у них переднего мозга, посылающего у полихет нервы к глоточному отделу. Сидячие полихеты, перешедшие к питанию грунтом или к седиментации наннопланктона, наряду с приобретением многих признаков специализации, утрачивают совершенство органов чувств и хищную глотку бродячих семейств; в связи с этим их головной мозг также значительно упрощается (рис. 96). Во внутреннем строении мозга полихет наиболее замечательной особенностью является возникновение стебельчатых тел (corpora pedunculata), являющихся высшими ассоциативными центрами в мозгу Articulata (рис. 95, Д). У сидячих полихет они редуцированы, у олигохет и пиявок отсутствуют. Стебельчатые тела полихет возникают в связи с нервами пальпов, обеспечивая их ассоциативные связи с другими частями мозга. Особенно сильно они развиты у компактных, активных, подвижных афродитид. Быстро прячущиеся в норку аннелиды содержат в своих двигательных центрах «гигантские» клетки, от которых отходят «гигантские» волокна, тянущиеся вдоль брюшной цепочки. Мы видели, что «гигантские» волокна имеются и у других удлиненных беспозвоночных, совершающих быстрые движения, как прячущиеся в норку Enteropneusta и Lingula, прыгающие десятиногие раки и саранчевые. Имеются они и у ланцетника. У личинки стрекозы Аnах гигантские волокна брюшной цепочки участвуют в рефлексе выбрасывания воды из прямой кишки. У Аnах и Sepia «гигантские» волокна являются лишь крайним «плюс-вариантом» в ряду изменчивости диаметров двигательных волокон. Полихеты имеют хорошо развитый глоточный нервный аппарат, ганглии которого развиваются за счет эпителия глотки и который соединяется с мозгом, с околоротовыми коннективами или с первыми брюшными ганглиями иногда несколькими парами корешков (рис. 95, Е). С буккальным нервным аппаратом моллюсков он, вероятно, гомологичен, хотя, может быть, и в самой общей форме, так же, как и со стоматогастрическим нервным аппаратом членистоногих. В некоторых случаях нервные клетки аннелид берут на себя инкреторную функцию. Так, в ганглиях брюшной цепочки многих полихет, пиявок и дождевых червей имеются двигательные клетки, посылающие свои отростки к мышцам кровеносных сосудов и одновременно выделяющие адреналин; другими словами, эти клетки воздействуют на кровеносный аппарат животного двояким путем – нервным и гуморальным. Далее, по-видимому, мозг полихет до надлежащей поры тормозит своим присутствием эпитокное преобразование задних сегментов; удаление средней части простомия Nereis вызывает эпитокную перестройку даже незрелых особей; по аналогии с насекомыми можно предположить выделение частью мозговых клеток специального гормона, тормозящего «метаморфоз». В последние годы, в мозгу и брюшной цепочке нереид, а также ряда сидячих полихет, олигохет и сипункулид доказано присутствие невро-инкреторных клеток. Для нереид доказано совпадение их циклов секреции с половыми циклами. У Nephthys, нереид, дождевых червей аксоны невро-инкреторных клеток мозга в значительной своей части заканчиваются в кровяном синусе или сплетении кровеносных сосудов, подстилающем надглоточный ганглий. Этот цереброваскулярный комплекс является типичным неврогемальным органом. Очень важную, но очень трудную задачу представляет гомологизация различных отделов нервного аппарата полихет, с одной стороны, сколецид и моллюсков – с другой. Говоря в самой общей форме, нервный аппарат высших турбеллярий состоит из кожного сплетения (ортогона) и церебрального ганглия (эндона), нервный аппарат трохофоры Lopadorhynchus – из нервного сплетения, соответствующего ортогону турбеллярий, и из циркумбластопорального сплетения, отсутствующего у этих последних. Часть ортогона личинки, лежащая в ее верхнем полушарии, дает начало головному мозгу взрослой полихеты; часть его, переходящая в нижнее полушарие, дает окологлоточные коннективы, а за счет циркумбластопорального сплетения образуются брюшные стволы. Таким образом, в состав мозга полихет входят части нервного аппарата личинки, отвечающие передней части ортогона турбеллярий. Но имеется ли в составе мозга полихет что-либо гомологичное эндону турбеллярий? Как мы видели, этот последний возникает в связи с аборальным органом чувств низших турбеллярий, статоцистом, и в соответствии с этим лежит на морфологической оси тела. У ряда полихет, например у Polygordius, в число зачатков, дающих начало мозгу, входят и элементы, связанные первоначально с теменной пластинкой, являющейся аборальным органом чувств трохофоры. Как легко видеть, эти элементы в какой-то мере являются гомологами эндона турбеллярий; однако в мозгу полихет они играют лишь незначительную роль. Тем не менее, теменная пластинка в качестве одного из зачатков мозга представляет значительное постоянство и, кроме полихет, прослеживается у моллюсков и даже у низших раков.
Центральный отдел нервного аппарата Onychophora и членистоногих состоит, как и у аннелид, из мозга, окологлоточных коннективов и брюшных стволов. Однако, как мы видели, в состав мозга представителей обоих подтипов входит по меньшей мере одна пара брюшных ганглиев, образующая тритоцеребральный отдел мозга. В силу этого мозг высших Articulata всегда лежит глубоко внутри тела и никогда не занимает у некоторых аннелид. Однако брюшная нервная лестница Notostraca лежит кнаружи от хорошо выраженного у них слоя продольных мышц, т.е. занимает более поверхностное положение, чем у многих полихет. Брюшные стволы всех членистоногих никогда не соединяются над анальным отверстием и почти всегда полностью ганглионизованы, причем в каждом сегменте обычно образуется по одной паре ганглиев. Однако у Scutigeromorpha ганглии брюшной цепочки не вполне обособлены от продольных коннективов, которые тоже покрыты клетками. Брюшная нервная лестница с широко расставленными стволами и длинными, притом двойными, поперечными комиссурами из всех современных членистоногих свойственна листоногим ракам (рис. 97, А) и Gephalocarida. У всех других членистоногих оба ганглия одного сегмента сближаются до соприкосновения, так что поперечная комиссура внешне становится невидимой, хотя коннективы очень часто остаются разделенными (большинство многоножек и ракообразных, часть насекомых). У многих раков, насекомых и всех наземных хелицеровых происходит, по крайней мере по внешности, полное слияние не только ганглиев, но и коннективов, в одну непарную цепочку. В этом отношении процессы централизации протекают у аннелид и членистоногих параллельно и в различных группах членистоногих независимо. Однако, в отличие от аннелид, у членистоногих широкое развитие получает также и концентрация брюшной цепочки путем укорочения продольных коннективов и слияния последовательных метамерных ганглиев. Явление это вполне аналогично такой же концентрации ганглиев моллюсков, но вследствие иного общего плана нервного аппарата протекает у членистоногих несколько иначе. В отдельных группах членистоногих характер и степень концентрации брюшной цепочки тесно связаны с характером и степенью гетерономности их метамерии, с объемом и степенью интеграции тагм, длиною и степенью интеграции всего тела. Нервный аппарат Phyllopoda представляет прототип гомономной брюшной лестницы, состоящей из независимых друг от друга ганглиев. В том числе независимы от мозга и тритоцеребральные ганглии. У всех остальных членистоногих, в связи с образованием головы, тритоцеребральные ганглии сливаются с мозгом. Но и все ганглии, кроме тритоцеребральных, остаются независимыми между собой лишь у немногих ракообразных. У громадного большинства ракообразных и Atelocerata сливаются между собой по меньшей мере ганглии мандибул и двух пар максилл в один подглоточный ганглий, однако у Chilopoda Аnаmorpha ганглии максилл II еще не вошли в его состав (рис. 98, Б). У всех насекомых и Chilopoda ганглии последних сегментов тела (9–11-й сегменты брюшка у насекомых, половые сегменты у Chilopoda) сливаются с ганглием предшествующего сегмента: 8-го брюшного сегмента у насекомых, сегмента последних бегательных ног у Chilopoda (рис. 98, Г). Дальнейшее слияние идет довольно разнообразными путями. У Crustacea Malacostraca прежде всего сливаются с подглоточным ганглием ганглии максиллипед (Decapoda Macrura, рис. 97, Е), затем другие грудные ганглии и, наконец, у крабов (Вrаchiura, рис. 97, Н) все ганглии брюшной цепочки сливаются в одну общую массу. Столь же высокая концентрация брюшной цепочки наблюдается и у некоторых Entomostraca (часть Сорepoda и Cirripedia), у которых, впрочем, она облегчается уменьшением числа сегментов тела. Во всех этих случаях централизация нервного аппарата достигает почти такой же степени, как и у заднежаберных улиток или головоногих. Тогда как у полихеты или листоногого рака каждый ганглий лежит в том сегменте тела, который он иннервирует, у краба все они сдвинуты в передние сегменты гнатоторакса, но каждый сохраняет связь со своей областью иннервации, каждый продолжает иннервировать свой сегмент. Таким образом, коннективы укорачиваются в той же мере, в какой удлиняются периферические нервы, и достигается основная задача централизации нервного аппарата – укорочение межцентральных путей.
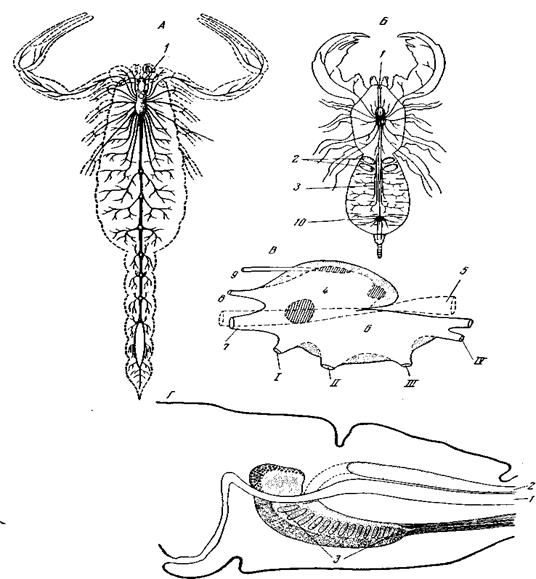
Рис. 97. Типы строения центрального нервного аппарата ракообразных А – Phyllopoda; Б – Cirripedia Pedunculata; В – Girripedia Operculata; Г – Euphausiacea; Тот же процесс концентрации брюшной цепочки независимым путем или, вернее, множеством независимых путей, протекает и в других классах членистоногих – у насекомых, хелицеровых и пантопод. Исключение составляют только многоножки, у которых в связи с незначительной интеграцией их туловища концентрация брюшной цепочки также останавливается на одной из первоначальных, вышеупомянутых ступеней развития (рис. 98, А). Среди насекомых даже наиболее примитивные формы, вроде Machilis (Thysanura, рис. 98, Д), имеют во взрослом виде уже лишь 8 свободных абдоминальных ганглиев. В большинстве случаев такое число ганглиев свойственно только личинкам. Почти все взрослые насекомые представляют более значительные степени концентрации брюшной цепочки. Прежде всего первый брюшной ганглий очень часто сливается с третьим грудным, и притом даже не только у тех насекомых, у которых сливаются соответствующие сегменты. У некоторых бабочек и прямокрылых с третьим грудным сливается не 1, а 2 первые брюшные ганглия (например, у Telea, сем. Saturniidae), у Locusta – целых три. Далее происходит слияние грудных ганглиев между собой и с подглоточным ганглием, а брюшных – между собой. И, наконец, у некоторых форм происходит, как у крабов, полное слияние всех ганглиев брюшной цепочки в одну общую подглоточную массу. Разнообразие сочетаний в отдельных случаях очень велико. Общее разнообразие еще усиливается наличием в некоторых случаях полового диморфизма, а у Holometabola – различиями между окрыленными насекомыми и личинками. Пользуясь главным образом данными Е. Brandt, изучившего анатомическое строение нервного аппарата очень многих насекомых, а также и другими источниками, можно составить следующую таблицу. Рассматривая эту таблицу, мы можем заметить несколько общих закономерностей. 1. Максимальной концентрации ганглиев головы и груди всегда соответствует высокая концентрация ганглиев брюшка. В силу этого в первых рядах отсутствуют наименее централизованные формы нервной цепочки брюшка. 2. Большинство Diptera располагается во втором ряду и очень немногие остаются за пределами этого ряда, т.е. для двукрылых характерно большое постоянство в числе ганглиев груди и головы при последователь-лом сокращении числа свободных ганглиев брюшка. 3. Все клопы оказываются в нулевом столбце, т.е. у всех Heteroptera ганглии брюшка слиты с последними грудными (рис. 99, 3), тогда как ганглии головы и груди представляют ряд последовательных стадий слияния между собой. Клопы – группа палеонтологически довольно древняя однако во многих отношениях высоко специализованная. Чертой специализации является у них и полная концентрация брюшной цепочки. Высокая степень концентрации брюшной цепочки свойственна также Homo-ptera и другим отрядам (Thysanoptera, Psocoptera, Anoplura). РАЗЛИЧНЫЕ СТУПЕНИ КОНЦЕНТРАЦИИ БРЮШНОЙ НЕРВНОЙ ЦЕПОЧКИ У НАСЕКОМЫХ (зависимость между числом свободных ганглиев в брюшке и в голове и груди)
Примечание. В таблице приведено число свободных ганглиев головы, груди и брюшка у различных насекомых. Значение сочетаний числа ганглиев головы и груди следующее: 1+1 означает 1 ганглий (надглоточный) в голове и в груди (последний охватывает подглоточный и все грудные ганглии и всю брюшную цепочку); 2+1 означает надглоточный, подглоточный и один, общий грудной; 2+2 означает надглоточный, подглоточный и 2 грудных; 2+3 надглоточный, подглоточный и 3 самостоятельных грудных
Рис. 98. Центральный отдел нервного аппарата низших членистоногих А – Polydesmus verhoeffi (Diplopoda): 1 – нерв антенн; 2 – надглоточный ганглий; 3 – подглоточный ганглий; 4 – глоточные коннективы (по Newport); Б – Thereuopoda clunifera (Chilopoda Scutigeromorplia), мозг и передняя часть брюшной цепочки, вид с брюшной стороны; обратить внимание на примитивные черты: отсутствие слияния ганглиев мандибул и максилл в один подглоточный ганглий и слабую ганглионизацию брюшной цепочки; В – Scolioplanes hirtipes (Chilopoda, Geophilomorpha), мозг и передняя часть цепочки, вид с брюшной стороны; обратить внимание на редукцию зрительных долей и всего протоцеребра под влиянием слепоты, связанной с роющим образом жизни, при общем более высоком уровне строения брюшной цепочки: 1 – предлобное сплетение, дающее нервы наличнику (clypeus) и верхней губе; 2 – мозговая железа; 3 – зрительные доли; 4 – тритоцеребрум; 5 – дейтоцеребрум; 6 – протоцеребрум; 7 – фронтальный ганглии; 8 – подглоточный ганглий; 9 – ганглий сегмента макслиллипед; мд – нервы мандибул; мкс 1 – нервы максилл первых; мкс II – нервы максилл вторых; мксп – нервы максиллипед. Нервы, отходящие от протоцеребра: н1 – к сложным глазам; н3 – к органу Томешвари; н4 – к мозговой железе. Нервы, отходящие от дейтоцеребра: н7 – чувствительный нерв антенн; н9- н11 – двигательные антенн; н12 – двигательный к некоторым мышцам головы; н13 – коннективы к фронтальному ганглию; н14 – возвратный нерв (n. recurrens), по ходу его вздутия – стоматогастрический ганглий; н15 – двигательный к глотке; н16 – корни непарного нерва, сопровождающего аорту; н17 – лабральные к предлобному сплетению; н18 – чувствительные к боковым частям ротовой полости; н20 – тритоцеребральная комиссура; н21 – к ретрактору языка; Г – Thereuopoda clunifera, задний конец брюшной нервной цепочки: 1 – ганглий 21-го сегмента; 2 – концевой ганглий, продукт слияния ганглиев генитального и прегенитального сегментов; нервы: 3 – к ректальному сплетению; 4 – к половым протокам; 5 – к париетальной мускулатуре; 6 – к гоноподиям; 7 – к последней паре бегательных ног (по К. Fahlander); Д – Machilis (Insecta, Thysanura), надглоточный ганглий и брюшная цепочка: 1— нервы к антеннам; 2 – надглоточный ганглий; 3 – подглоточный ганглий; 4 – зрительные доли; 5 – непарный медиальный (Заварзинский) нерв; 6 – пищевод; I–III – ганглии груди; IV–XI – ганглии брюшка (последний из них состоит из трех слившихся ганглиев) (по Oudemans) 4. Среди Coleoptera и Diptera высшие представители (Lamellicornia среди жуков, Muscoidea среди двукрылых) обладают наиболее концентрированной нервной цепочкой, наиболее примитивные (Diptera Nematocera, жуки Cicindela, Dictyopterus) наименее концентрированной (рис. 99, А–Ж). 5. Бабочки почти все сосредоточены в четвертом столбце третьего и четвертого рядов, т.е. обладают четырьмя ганглиями брюшка и только Hepialus, форма очень примитивная и в других отношениях, попадает в пятый столбец, так как имеет пять ганглиев брюшка; у самки чехлоноски Pachytelia сохраняется гусеничное число брюшных ганглиев, однако эта форма вообще сильно упрощенная, с рядом неотенических признаков. 6. Представители примитивных отрядов (Protura, Thysanura, Odona-ta, ортоптероидные отряды, Neuroptera), как правило, обладают весьма низкой степенью концентрации брюшной цепочки, но специализованные формы среди низших отрядов могут попадать и в нулевой столбец, как, например, Podura (Сollembola). Таким образом, таблица в целом с несомненностью показывает, что у насекомых процесс концентрации брюшной цепочки происходит независимо в различных отрядах, а иногда даже и в различных семействах одного и того же отряда (Coleoptera).
Рис. 99. Схема строения центрального нервного аппарата насекомых A – Dictyopterus sauguineus; Б – Cicindela sylvatica; В – Gyrinus natator; Г – Rhizotrogus solsticialis (вес четыре – Coleoptera); Д – Chironomus plumosus; Е – Tabanus bovinus;
Несколько слов относительно развития брюшной цепочки. У зародышей насекомых все ее ганглии независимы друг от друга. У личинки, как общее правило, цепочка концентрирована больше, чем у зародыша, у взрослого насекомого – еще больше (рис.100, А, Б). Но из этого правила существует несколько интересных исключений, особенно среди личинок мух. Так, у личинки Syrphus или Volucella, по Брандту, брюшная цепочка полностью концентрирована, тогда как у взрослых сирфид в брюшке имеются два свободных ганглия (рис. 100, В, Г). Другими словами, с развитием здесь происходит децентрализация. Этот случай представляет один из примеров биогенетического «закона» наизнанку: личинка сирфа в строении своей брюшной цепочки как бы забегает вперед в общем направлении эволюции данной группы, а имагинальная форма отстает. Понятно, что отдельные метамерные компоненты концентрированного брюшного мозга личинки мухи посылают периферические нервы к своим сегментам (закон сохранения области иннервации), но они получают и трахейные веточки от метамерных веток своих сегментов (рис. 100, Д). Сохранение области иннервации или области питания есть своего рода инерция, и в дальнейшем ходе эволюции вполне возможны перестройки.
Рис. 100. Развитие брюшной нервной цепочки у насекомых Нервный аппарат: А – гусеницы-капустницы Pieris brassicae (Lcpidoptera); Б – взрослой бабочки того же вида (по Брандту); В – личинки Volucella zonaria (Diptera); Г – взрослой мухи (по Kunckel d'Herculais); Д – личинки Phormia regina (Diptera), изображены метамерные нервы, идущие от лежащих в груди брюшных ганглиев к своим сегментам, и метамериые ветви трахей, идущие к тем же сдвинутым вперед и слитым в одну массу ганглиям: 1 – переднее дыхальце; В классе Chelicerata мы также наблюдаем постепенную концентрацию брюшной цепочки, однако ранних стадий этого процесса мы здесь уже не застаем. Наименее концентрирована брюшная цепочка у мечехвоста; однако и у него все ганглии просомы слиты воедино вблизи пищевода; зато все ганглии мезосомы остаются независимыми и имеется отдельный ганглий для трех слитых вместе сегментов метасомы. Кроме того, оба брюшных ствола личинки мечехвоста довольно далеко расставлены друг от друга, напоминая в этом отношении Phyllopoda. У скорпионов (рис. 101, А) все ганглии просомы и 4 ганглия мезосомы слиты в одну подглоточную массу, за которой идет цепочка из 7 ганглиев, иннервирующих два сегмента мезосомы и 6 сегментов метасомы. Последний ганглий метасомы, залегающий в 5-м ее сегменте, иннервирует также 6-й сегмент и тельсон. У Telyphones (рис. 101, Б) все ганглии брюшной цепочки слиты воедино, и лишь один из них, тройной по своему происхождению, вынесен далеко кзади (так же как у Stylops из отряда Strepsiptera среди насекомых) и иннервирует три последних сегмента тела. Такое же строение сохраняется у бихорок (Solifugae). У Araneina весь центральный отдел нервного аппарата, включая мозг и брюшные стволы, сливается в одну сплошную массу, пронизанную пищеводом. Однако у Mesothelae (= Liphistiomorpha) в ней хорошо различимы 17 невромер (рис. 101, Г), отвечающих полному числу постхелицеральных сегментов. У остальных Araneina закладывается столько же невромер, но 5 задних дегенерируют и в состав брюшной нервной цепочки входят только 12. Так же слиты в одну массу все ганглии тела и у лжескорпионов (рис. 101, В), сенокосцев и у всех изученных в этом отношении клещей. Из всего сказанного видно, что среди Chelicerata, так же как и в других классах, существует корреляция между степенью укорочения и интеграции тела с одной стороны, и степенью концентрации брюшной цепочки – с другой. Кроме того, мы еще видим, что ганглии просомы сливаются между собой первыми, ганглии мезосомы – за ними, а ганглии метасомы почти всюду, где сохраняются сегменты метасомы, обладают частичной независимостью, и полная концентрация брюшной цепочки обычно достигается лишь после полной редукции метасомы. Однако при высоко интегрированной форме тела полная концентрация брюшной цепочки встречается и при наличии остатков метасомы, как, например, у лжескорпионов. Итак, среди Chelicerata многие отряды достигают такой же или еще более высокой концентрации нервного аппарата, какой среди ракообразных достигают крабы и некоторые специализованные группы Entomost-rаса, среди насекомых – Hydrometra или Rhizotrogus и среди аннелид –
Рис. 101. Схемы центрального нервного аппарата паукообразных А – Androctonus (Scorpionoidea); Б – Telyphonus caudatus, оба с дорсальной стороны;
Во внутреннем строении мозга отметим только некоторые черты. Для большинства членистоногих характерно мощное развитие и громадная роль зрительных центров, что, несомненно, связано с развитием сложных глаз. Зрительные центры входят в состав протоцеребра и здесь же располагаются стебельчатые тела, также достигающие у членистоногих высшей степени развития. Стебельчатые тела членистоногих связаны как со зрительными, так и с обонятельными центрами (антеннальные гломерулы), расположенными в дейтоцеребре. Можно сказать, что первоначальное развитие мозга членистоногих идет под влиянием двух главных органов чувств – глаз и антенн. У хелицеровых вместе с антеннами исчезает дейтоцеребрум. У Chilopoda Epimorpha параллельно с редукцией глаз идет и редукция зрительных долей переднего мозга, наиболее полная у вполне безглазых Geophilomorpha (рис. 98, В). Вообще сильная редукция органов чувств, там, где она имеет место, обычно вызывает у членистоногих значительные упрощения в строении мозга, что мы наблюдаем у большинства паукообразных, у пантопод, у паразитических ракообразных и других.
Рис. 102. Усиление стебельчатых тел мозга с осложнением поведения у перепончатокрылых A – мозг Tenthredo flava (пилильщик), вид спереди; Б – Ichneumon obsessor (наездник); В – Andrena albicans (одиночная пчела); Г – Vespa vulgaris (общественная оса); 1 – лобные глазки; 2 – зрительные доли; 3 – сетчатка сложного глаза; 4 – дейтоцеребрум; По внутреннему своему строению мозг всех членистоногих представляет видоизменения одного общего плана, причем ракообразные, многоножки и насекомые и в этом отношении стоят ближе друг к другу, нежели те и другие к Chelicerata. Детальное сравнение структуры мозга насекомых с таковой ракообразных и многоножек позволяет сделать вывод, что по основным организационным признакам мозг насекомых несколько ближе к таковому многоножек, тогда как по уровню дифференциации своего мозга высшие насекомые очень близки к десятиногим ракам, а многоножки и низшие насекомые, например Campodeoidea, стоят приблизительно на уровне Phyllopoda. При этом направление развития в обоих стволах очень сходно; особенно велико сходство между насекомыми и десятиногими раками в строении сложных глаз и зрительных центров. Степень развития стебельчатых тел у различных членистоногих представляет некоторые замечательные закономерности, заставляющие думать, что эти образования являются высшими центрами, обеспечивающими возможность наиболее сложных форм поведения, свойственных членистоногим. Действительно, ряд данных указывает, что качественное и количественное развитие этих органов стоит в прямой корреляции со сложностью поведения животного. Среди перепончатокрылых наименее развиты стебельчатые тела у пилильщиков (Tenthredinidae, рис. 102, А) и рогохвостов (Siricidae), имеющих наиболее простые материнские инстинкты, лучше развиты они у наездников (Ichneumonidae, рис. 102, Б), еще лучше – у одиночных пчел (Andraenа, рис. 102, В; Anthophora, и лучше всего – у общественных ос (Vespa, рис. 102, Г), обладающих из всех изученных им насекомых наиболее сложными инстинктами. Среди пчел наибольшего развития стебельчатые тела достигают у общественных форм, Apis и Bombus. При этом, в соответствии с их ролью в жизни вида, самки шмелей имеют стебельчатые тела, развитые не хуже, чем у рабочих, тогда как у домашней пчелы рабочие бесспорно стоят на первом месте. У муравья Camponotus, зрительные центры развиты лучше всего у самцов, затем у самок, затем у рабочих, развитие же обонятельных центров и стебельчатых тел стоит в обратном порядке: объемы стебельчатых тел у рабочих, самок и самцов этого муравья относятся друг к другу, как 8:4:1. По относительным размерам своих стебельчатых тел рабочие муравьи занимают среди насекомых первое место. Наряду с указанной корреляцией существует другая: чем больше размеры тела какого-либо членистоногого, тем сильнее, при прочих равных условиях, развиты его стебельчатые тела. Из всех изученных членистоногих сильнее всего они развиты у Limulus. У крупной Vespa crabro они развиты сильнее, чем у более мелкой V. vulgaris. Очень крупных размеров достигают они у десятиногих раков и у некоторых из телифонов. При этом ни Limulus, ни телифоны не отличаются сложностью поведения. На основании этих данных некоторые авторы возражали против значения стебельчатых тел, как центров высшей нервной деятельности. Однако возражения эти вряд ли справедливы, как легко видеть из аналогии с млекопитающими, у которых степень развития извилин коры большого мозга также связана не только со сложностью поведения того или иного животного, но и с размерами его тела.
Поиск по сайту: |