|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
КЛАСС П. БРЮХОНОГИЕ (GASTROPODA)
Брюхоногие, или улитки, — самый богатый представителями класс моллюсков.
Первично брюхоногие моллюски — обитатели моря, но многие из них приспособились к жизни в пресных водоемах и на суше. Очень небольшое число видов ведет паразитический образ жизни. Класс Gastropoda подразделяется на 3 подкласса: пе-реднежаберных (Prosobranchia), заднежаберных (Opisthobranchia) и легочных (Pulmonata). Размеры брюхоногих варьируют от 2—3 мм до нескольких десятков сантиметров. К самым крупным видам относятся Hemifusus proboscidiferus (подкл. Prosobranchia) с раковиной в 60 см, морской заяц — Aplysia (подкл. Opisthobranchia), достигающий 25 см, и некоторые африканские виды наземных улиток Achatina (подкл. Pulmonata), плавающая форма Pte-rotrachea coronata, длина которой превышает 25 см, и др.
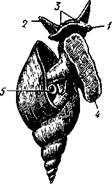
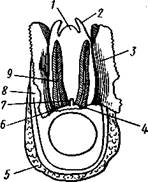
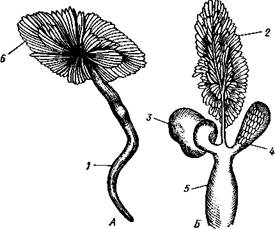
Голова брюхоногих явственно обособлена от тела, нога хорошо развита и обычно имеет широкую полза-тельную подошву (рис. 438), туловище образует кверху вырост в виде большого внутренностного мешка. Раковина состоит из одного куска, но иногда может подвергаться редукции. Характерная черта всех брюхоногих— асимметричность строения. Обыкновенно она уже с первого взгляда бросается в глаза. Асимметрия брюхоногих моллюсков выражается, во-первых, в редукции правых органов мантийного комплекса, сопровождающейся усиленным развитием их партнеров на левой стороне, и, во-вторых, внутренностный мешок закручивается спирально — эта особенность внешне ясно выражена в форме раковины (рис. 438). Строение и физиология. Форма тела брюхоногих разнообразна, но большей частью тело продолговато и на спинной стороне выпукло. Голова хорошо развита. Она несет рот, 1—2 пары щупалец и пару глаз. Передняя часть головы нередко вытягивается в длинный хобот, на конце которого располагается ротовое отверстие. Нога представляет мускулистый брюшной вырост с плоской полза-тельной подошвой; при помощи сокращений ноги животное медленно и плавно ползет по субстрату. В зависимости от образа жизни нога можгт испытывать различные модификации. Сильнее всего изменена нога у не-' которых представителей кл. Gastropoda, перешедших к плавающему образу жизни. Так, у киленогих (подкл. Prosobranchia, с. 471) нога сплющивается с боков и превращается в узкий вертикальный плавник (рис. 439), иногда еще с маленьким рудиментом подошвы на брюшном крае, которым животное оживленно машет в обе стороны. В то же время задний отдел ноги вытягивается в длинный хвостовидный руль. У другой группы плавающих Gastropoda, а именно у крылоногих (подкл. заднежаберных— Opisthobranchia, с. 472), боковые части ноги сильно разра- стаются, образуя пару бьющих по воде крыловидных лопастей (рис. 440), тогда как основная центральная часть редуцируется. Туловище, или внутренностный мешок, у низших брюхоногих (например, у некоторых Diotocardia из подкл. Prosobranchia) иногда симметричен и сравнительно неясно отграничен от ноги, так же как и у голожаберных (Nudibranchia из подкл. Opisthobranchia, с. 472) и у некоторых
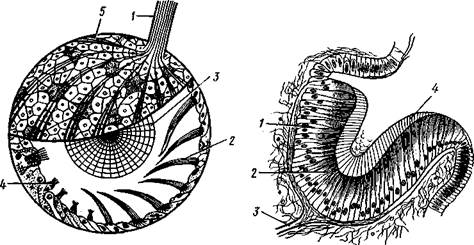
Рис. 439. Киленогий моллюск Pterotrachea coronata (из Каруса): 1 — передняя часть головы, 2 — глотка, 3 — статоцист, 4 — головная артерия, 5 — ножная артерия, 6" — передняя часть ноги, 7 — хвостовой придаток, 8 — задняя часть ноги, 9 — ктенчдий. 10 — анальное отверстие, 11 — предсердие, 12 — желудочек сердца, 13 — осфрадий, 14 — висцеральный ганглий, 15 — кишка, 16—педальный ганглий, 17—церебральный ганглий, 18 — глаза легочных (подкл. Pulmonata). У большинства видов класса Gastropoda туловище, напротив, выдается над ногой в виде большого, более или менее спирально закрученного мешка. На туловище образуется по направлению книзу складка покровов, мантийная складка, под которой находится мантийная полость с расположенным в ней мантийным комплексом органов (см. 442). Как складка, так и полость обычно развиты на передней и на правой стороне туловища. Мантия выделяет раковину, в большинстве случаев закрученную спирально. На начальном конце раковина слепо замкнута — это ее вершина, на другом конце имеется отверстие— устье, через которое торчат наружу голова и нога животного. Лишь в редких случаях обороты раковины лежат в одной плоскости (например, у примитивной ископаемой формы Tremonotus, см. рис. 457, или у современных брюхоногих, например у катушки —Planorbis). Обычно спираль коническая (турбоспиралъ). Диаметр оборотов в соответствии с ростом животного увеличивается от вершины к устью. При этом каждый новый, более крупный оборот раковины может охватывать все предыдущие, делая их незаметными (Сургаеа); такие раковины называют инволютными. Гораздо чаще последний оборот лишь прилегает к предыдущим, так что все они заметны. Среди брюхоногих со спиральными раковинами различают два типа по направлению закручивания спирали. Если при взгляде со стороны вершины спираль закручивается (начиная от вершины) по направлению движения часовой стрелки, то раковина называется правовращающей или дексиотропной — таких преобладающее большинство. Если же она закручивается влево, то мы имеем дело с левовращающей, лейотропной раковиной. Ось, вокруг которой закручена спираль, обыкновенно представлена плотным столбиком— колонкой (рис. 441); если же обороты не соприкасаются своими внутренними стенками, то вместо плотной колонки по оси спирали обра- зуется полость, пупок. Спиральная раковина обычно настолько обширна, что при опасности животное целиком может втягиваться в нее. У ряда форм, главным образом относящихся к подкл. Prosobranchia, на спинной стороне задней части ноги обычно имеется известковая или роговая пластинка, крышечка, более или менее соразмерная с диаметром
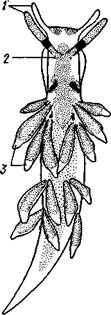
Рис. 440. Крылоногий моллюск Clione limacina (по Боасу): ; — околоротовые придатки, 2 — щупальца, 3 — плавник — боковая часть ноги, 4 — совокупительный орган, 5 — внутренностный мешок (туловище), 6 — средняя часть ноги Рис. 441. Продольный распил раковины Cha- ronia (по Оуэну): 1 — вершина раковины, 2 — столбик, 3 — сифо-нальный вырост раковины устья, для замыкания которого она и служит при втягивании животного. Некоторые виды брюхоногих имеют более просто устроенную раковину колпачковидной формы (см. рис. 462, Г). Это не примитивные, но вторично упрощенные в отношении раковины брюхоногие, так как личинки их сначала обладают более или менее завитой раковиной, которая лишь позже принимает вид колпачка. Раковина брюхоногих состоит из тонкого органического наружного слоя (periostracum), под которым лежит фарфоровидвый слой (ostra-cum), образованный известковыми пластинками, расположенными перпендикулярно к поверхности раковины. Углекислая известь добывается из заглатываемой животным воды и пищи. У некоторых брюхоногих (Haliotis, Turbo и др.) имеется еще внутренний перламутровый слой (с. 476).
Во всех подклассах брюхоногих нередко наблюдается редукция ра
ся и отчасти прикрывается двумя заворачивающимися на нее складками мягких тканей. В одних случаях это могут быть складки мантии, в других к ним добавляются разрастающиеся боковые участки ноги; 3) складки срастаются друг с другом по срединной линии над раковиной (p. Aplysia, слизень Limax) — раковина из наружной становится внутренней и превращается в тонкую пластинку; 4) рудиментарная раковина распадается на отдельные известковые тельца, лежащие в покровах спины — слизень Arion (рис. 442); голожаберные, или Nudibranchia (рис. 443); 5) все следы раковины пропадают — Pterotrachea (см. рис. 439). Утрата раковины чаще всего наблюдается у плавающих и наземных форм, где польза раковины как защитного приспособления, очевидно, меньше невыгодной и при плавании и ползании ее тяжести. Исчезает раковина и у паразитических брюхоногих.
Пищеварительная система. Рот лежит на нижней стороне переднего конца головы, которая у некоторых форм вытягивается и образует так называемое рыло. Еще более удлиняясь, передний конец головы превращается у некоторых хищных форм из подкл. Prosobranchia в мускулистый хоботок, который может вворачиваться и выбрасываться наружу при захватывании добычи. Рот ведет в ротовую полость, переходящую в мускулистую глотку (рис. 444). Глотка содержит язык — мускулистый валик, покрытый тонкой кутикулой и несущий твердые зубцы, расположенные поперечными рядами (рис. 445). Кроме того, в ротовой полости на границе с глоткой лежат так называемые челюсти — местные утолщения кутикулы; они роговые, а иногда содержат отложения извести. Число и расположение челюстей варьируют. В глотку открываются протоки одной пары слюнных желез. Слюнные железы некоторых хищных переднежаберных (Топпа, Cassis и др.) замечательны большим содержанием в их секрете свободной серной кислоты (крепость до 4%)- Кислый секрет служит этим хищникам для растворения раковины других моллюсков или панцирей иглокожих, которыми они питаются. Глотка переходит в довольно длинный пищевод, образующий у некоторых брюхоногих местное расширение, или зоб (рис. 446). Все описанные до сих пор части принадлежат эктодермальной передней кишке. Начальная часть энтодермальной средней кишки образует мешковид-ный желудок, в который впадают протоки «печени». Печень закладывается в виде парного выпячивания кишечника, но у взрослых брюхоногих в связи с их асимметрией во многих случаях представляет непарный орган; одна половина недоразвивается. Печень брюхоногих объемиста и состоит из многочисленных долек, протоки которых соединяются вместе, а затем впадают в желудок. Секрет печени растворяет углеводы, но, кроме того, печень обладает способностью к всасыванию пищи (подобно тонкой кишке позвоночных), а также служит для отложения в ней жира, гликогена. Часто, особенно у низших брюхоногих, пища подвергается в желудке сортировке, и более мелкие частицы мерцанием ресничного эпителия желудка направляются через протоки печени в полости печеночных долек. Здесь они захватываются клетками и перевариваются
внутриклеточно (фагоцитоз). В желудке некоторых низших брюхоногих имеется хрустальный столбик — студенистое образование, состоящее из белков и связанных с ними ферментов — амилазы и целлюлазы. Конец столбика в щелочной среде желудка постепенно растворяется, освобождая ферменты, способствующие перевариванию углеводов пищи.
Рис. 444. Срединный разрез через глотку виноградной улитки Helix pomatia (по Траппману): / — челюсть, 2 — ротовая полость, 3 — мускульная стенка глотки, 4 — кровеносные полости языка, 5 — радулярный хрящ, 6 — внутренняя глоточная мышца, 7 — складка глоточного эпителия, 8 — образующий радулу эпителий, 9 — радулярное влагалище, 10 — соединительная ткань радулярного влагалища, 11 — мускул, поддерживающий радулярное влагалище, 12 — пищевод, 13 — буккальная комиссура, 14 — полость глотки, /5 — радула, 16 — язык, 17 — Кутикула Интересные изменения испытывает печень у голожаберных (подкл. Opisthobran-chia, с. 472). У многих из них печень как бы разбивается на систему отходящих от кишечника кверху железистых каналов, веточки которых заходят в покрывающие спину щупальцевидные выросты — вторичные жабры и на вершине их могут даже сообщаться с наружной средой. Особенно интересно, что в эпителии этих веточек печени — там, где они сообщаются с внешней средой, на концах спинных жабр нередко наблюдается присутствие многочисленных стрекательных капсул (например, у сем. Aeolididae). Прежде считали, что стрекательные капсулы принадлежат самому организму голожаберных, но специальные эксперименты показали, что они попадают в моллюска из пищи. Именно представители сем. Aeolididae питаются гидроидными полипами, стрекательные капсулы которых не перевариваются моллюском, но поступают в печеночные выросты
Рис. 445. Часть радулы морского блюдечка PateUa роп-Нса (по Иванову) хищника, сохраняя там способность функционировать так же, как они функционировали в теле полипа. За желудком следует тонкая кишка, делающая одну или несколько петель, но затем заворачивающаяся кпереди и переходящая в заднюю кишку. Последняя заканчивается порошицей, лежащей или на переднем конце туловища, над головой, или где-нибудь на правой стороне тела.
Рис. 446. Вскрытая виноградная улитка (из Бриана): 1 — губное щупальце, 2 — глазное щупальце, 3 — глотка, 4 — церебральный ганглий, 5 — легкое, 6 — легочная вена, 7— перерезанное легочное отверстие, S — анальное отверстие, 9 — отверстие мочеточника, 10 — прямая кишка, II — мочеточник, 12 — предсердие, 13 — желудочек сердца, 14 — перикардий, 15 — почка, 16 — желудок, 17 — печень, 18 — гермафродитная железа, 19 — герма-фродитный проток, 20 — белковая железа, 21 — семяприемник, 22 — канал семяприемника, 23 — яйцесемяпровод. 24 — семяпровод, 25 — мешок любовных стрел, 26 — пальцевидные железы, 27 — бич, 28 — пенис, 29 — слюнные железы
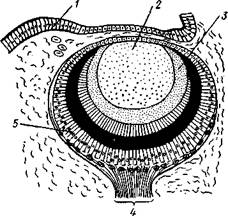
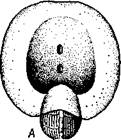
Рис. 447. Симметричный пе-реднежаберный моллюск Fissurella. Вид со спины, ра-копина удалена, мантийная полость вскрыта (по Пель-зенеру): / — голова, 2 — щупальца, 3 —• отвернутый лоскут мантии, 4 — отверстие правой почки, 5 — нога, 6 — отверстие левой почки, 7 — анальное отверстие, 8 — часть верхнего отверстия мантийной полости, 9 — левый кте-нидий
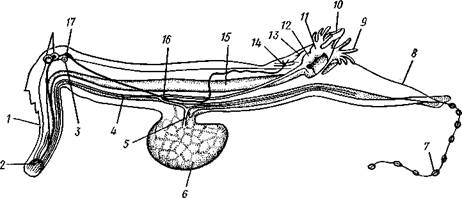
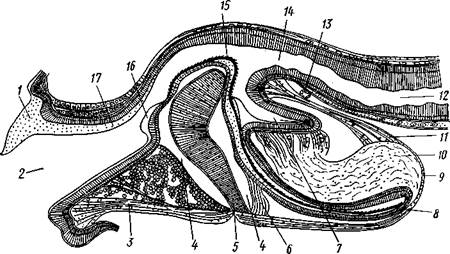
Интересно, что у некоторых низших брюхоногих задняя кишка проходит сквозь желудочек сердца. Органы дыхания. Большинство брюхоногих дышит жабрами. Первичными, или настоящими, жабрами являются ктенидии, парные органы, расположенные по бокам от порошицы. У многих форм они имеют вид вытянутых двоякоперистых придатков, заостряющихся к свободному концу. Каждый ктенидии состоит из осевого уплощенного стволика, несущего два ряда лепестков. Ктенидии характеризуются присутствием у их основания органов химического чувства — осфрадиев. Первично ктс-нидиев одна пара (рис. 447), но в связи с недоразвитием органов правой стороны тела у большинства брюхоногих правый ктенидии атрофируется. Так, у некоторых переднежаберных (Haliotis) он меньше левого. У высших представителей этой группы жабра уже одна, и притом нередко из двоякоперистой она становится однорядноперистой вследствие прирастания одной стороной к стенке мантии. Заднсжаберные (Opistho-branchia) в лучшем случае сохраняют один ктенидии, который часто сильно смещен кзади по правой стороне и смотрит концом назад, между тем как ктенидии Prosobranchia лежат ближе кпереди и смотрят концами вперед. В каждом подклассе Gastropoda есть формы, у которых настоящие жабры исчезли и вторично заменились другими органами дыхания. У водных брюхоногих в таком случае на самых различных местах тела могут возникать выросты, физиологически соответствующие ктенидиям, но не гомологичные им. Все эти образования называются вторичными или адаптивными жабрами (см. рис. 443). Наконец, у наземных легочных брюхоногих (подкл. Pulmonata) водное дыхание заменилось воздушным, ктенидии исчез и для дыхания служит легкое. Участок мантийной полости у них обособляется и открывается наружу самостоятельным отверстием (см. рис. 438). Это так называемая легочная полость, в стенках которой развиваются многочисленные кровеносные сосуды (см. рис. 446). Легкое сохраняется в виде единственного дыхательного органа у многих Pulmonata, несмотря на то что некоторые легочные моллюски вернулись к водному, а именно пресноводному образу жизни. Такие виды (прудовики, катушки и др.) дышат воздухом, периодически поднимаясь на поверхность воды. Кровеносная система (см. рис. 446). Положение центрального органа кровеносной системы — сердца — в теле брюхоногих моллюсков и особенности его строения неодинаковы в разных систематических группах. Наиболее примитивные формы подкласса Prosobranchia обладают симметричным сердцем, состоящим из желудочка и двух предсердий, расположенным по срединной линии тела на переднем его конце за головой; желудочек при этом пронизан задней частью кишечника. Однако по мере редукции правой жабры сердце также испытывает одностороннюю атрофию, выражающуюся в постепенном исчезновении правого предсердия. Так, у ряда видов переднежаберных правое предсердие уменьшено и слепо замкнуто, ибо ввиду редукции правой жабры исчезла и жаберная вена, впадающая в это предсердие. Вместе с тем несколько меняется и положение сердца: функционирующее предсердие ложится впереди желудочка. У всех прочих брюхоногих (высшие представители подкл. Prosobranchia, подкл. Opisthobranchia и Pulmonata) сохраняется только одно левое предсердие, тогда как правое полностью исчезает. Положение оставшегося предсердия зависит от положения жабры или легкого. У переднежаберных и легочных оно остается впереди желудочка, а у заднежа-берных ложится позади него, так как жабра их смещается далеко назад. Сердце обычно помещается над задней кишкой. Кроме того, оно всегда окружено перикардием, представляющим участок вторичной полости тела (целом).
Периферическая часть кровеносной системы в главных чертах дает следующую картину. От желудочка отходит аорта, которая вскоре делится на два ствола: головную аорту (к переднему концу тела) и внутренностную аорту (к кишечнику, печени и половой железе). Хорошо развитая артериальная система образована сосудами с обособленными стенками. У легочных моллюсков более крупные сосуды постепенно переходят в капилляры. В конце концов, однако, кровь изливается в мелкие лакуны соединительной ткани, теряет кислород и постепенно собирается в более крупные венозные лакуны, наиболее развитая из них омывает желудок, печень и гонаду. Отсюда кровь возвращается к органам дыхания (жабрам или легкому) и, окислившись, попадает непосредственно в сердце. Таким образом, сердце брюхоногих наполняется артериальной кровью. Кровь чаще всего бесцветна и содержит амебоциты. Иногда в крови имеется вещество, близкое к гемоглобину. У некоторых брюхоногих удалось обнаружить присутствие в крови белковых соединений, содержащих марганец, который у них играет, по-видимому, такую же роль, как железо в гемоглобине крови позвоночных. Нервная система брюхоногих (рис. 448) хорошо развита и представляет сравнительно-анатомический интерес. В общем, от низших брюхоногих к высшим наблюдается постепенный переход от системы, напоминающей таковую подтипа Amphineura, к разбросанно-узловой нервной системе из нескольких ганглиев, соединенных между собой свободными от нервных клеток волокнистыми перемычками. У наиболее примитивных брюхоногих моллюсков ганглии либо отсутствуют, либо выражены еще очень слабо. Нервные клетки в этом случае располагаются по ходу стволов. Последующие изменения происходят путем концентрации ганг-лиозных клеток в некоторых точках нервных стволов, в которых и образуются ганглии. У переднежаберных моллюсков первично обособляется пять пар основных ганглиев. 1. Над глоткой расположены два церебральных ганглия, соединенных церебральной комиссурой. 2. В ноге педальные стволы концентрируются в передней части в два педальных ганглия, соединенных под глоткой педальной комиссурой. Кроме того, они связаны с церебральными ганглиями двумя продольными перемычками, или коннективами (напомним, что поперечные нервные перемычки, соединяющие одноименные ганглии, принято называть ко-миссурами, продольные перемычки между ганглиями разного наименования— коннективами). У ряда примитивных современных форм обра-
зования педальных ганглиев еще не произошло, и вместо них сохраняются 2 педальных ствола. Далее на пути плевровисцеральных стволов обособились еще три пары ганглиев. 3. Два плевральных ганглия расположены приблизительно на уровне педальных ганглиев; они соединены коннективами с церебральными и педальными узлами. 4. Далее кзади на пути плевровисцеральных стволов залегают два париетальных ганглия. 5. Наконец, еще дальше, под задней кишкой, лежит пара висцеральных ганглиев, соединенных друг с другом висцеральной комиссурой. Последние три пары ганглиев как бы нанизаны на ту нервную петлю, которая образуется у подтипа Amphineura плевровисцеральными стволами. Участок стволов, находящийся между плевральным и висцеральным узлами каждой стороны и разбитый париетальным ганглием на 2 коннектива, называется плевровисцеральным коннективом. Кроме этих основных ганглиев образуются еще дополнительные ганг-лиозные массы, иннервирующие отдельные органы (например, глоточный, или буккальный ганглий; рис. 448). У заднежаберных (подкл. Opistho-branchia) и легочных (подкл. Pulfnonata) моллюсков первично обособляется не 5, а 7 пар ганглиев. У всех моллюсков эти цифры могут уменьшаться за счет слияния ганглиев в более сложные нервные узлы. Своеобразные изменения нервной системы брюхоногих моллюсков связаны с появлением у них асимметрии. Происхождение этих изменений подробнее будет рассматриваться дальше (с. 465). Главная суть их в следующем. У всех переднежаберных и немногих низших представителей двух других подклассов плевровисцеральные коннективы образуют перекрест так, что находящиеся на их протяжении париетальные ганглии перемещаются: левый на правую сторону (под кишку), а правый — на левую сторону тела (над кишечником). Нервную систему таких брюхоногих называют хиастоневральной или перекрещенной (рис. 448). У основной массы легочных и заднежаберных моллюсков перекрест коннективов вторично исчезает, но правый плевровисцеральный коннек-тив часто заметно короче левого. Это брюхоногие с неперекрещенной, или эутиневральной, системой. У высших Gastropoda плевровисцеральные коннективы вообще настолько укорачиваются, что ганглии собираются в одну общую массу вокруг глотки. Церебральными ганглиями иннервируются глаза, статоцисты, глотка и головные щупальца, педальными — мускулатура ноги. Плевральные ганглии снабжают нервами главным образом мантию. Париетальными ганглиями иннервируются ктенидии и осфрадии; наконец, висцеральными ганглиями осуществляется иннервация внутренностей. Органы чувств. Для осязания служат головные щупальца (см. рис. 438; 443), края мантии и некоторые другие места. Кроме того, имеются органы химического чувства. Ими считают прежде всего осфрадии, лежащие у основания ктенидиев. В наиболее развитом состоянии они имеют форму продолговатого валика, лежащего у основания жабры и .но обеим своим сторонам несущего 100—150 листочков. Вследствие этого осфрадии иногда очень похож по виду на жабру. Внутренность валика занята скоплением ганглиозных клеток, от которого отходят нервы к листочкам. Передняя пара головных щупалец, называемых губными, тоже, по-видимому, служит для восприятия химических раздражений, исполняя роль органов вкуса и обоняния. Органы равновесия в виде пары статоцистов имеются у всех Gastropoda. Это обыкновенно два замкнутых пузырька, эпителий которых состоит из мерцательных и чувствительных клеток (рис. 449); последние иногда образуют на стенке пузырька особое скопление — «слуховое пятно». В жидкости, заполняющей пузырек, плавают от 1 до 100 конкре-
Рис.449. Статоцист киленогого моллюска Pterotrachea (по Бючли): / — нерв. 2 —пучок ресничек, 3 — статолит, 4 — чувствительные клетки, 5 — нервные волокна Рис. 450. Разрез глаза морского блюдечка Patella rota (по Гильгер): J — сетчатка, 2—пигментированная зона, 3 —• зрительный нерв, 4 — утолщенная кутикула ций углекислой извести — статолитов, которые играют роль слуховых камешков. Различное положение их в статоцистах и надавливание то на одни, то на другие чувствительные клетки позволяют животному ориентироваться в пространстве. Оба статоциста лежат всегда в тесном
соседстве с педальными ганглиями, но иннервируются они тем не менее от церебральных ганглиев при помощи двух нервов. Одна пара глаз имеется почти у всех брюхоногих; они лежат на голове у основания, а иногда на вершине задней пары щупалец, соответственно часто обозначаемых как глазные. Сложность строения глаз варьирует от поо-стых ямок до глазных пузырей с хрусталиком и стекловидным телом (рис. 450, 451).
Выделительная система брюхоногих состоит из пары почек типа целомодуктов, из которых Рис. 451. Разрез глаза Fissurclla (из Догеля): / — кожный эпителий головы, 2 — хрусталик, 3 —стекловидное тело, 4 —зрительный нерв, 5 — зрительные клетки (сетчатка)
т. е. с участком целома, другим — открываются в мантийную полость сбоку от порошицы. Две почки имеются только у низших представителей подкл. Prosobranchia, но одна из них развита слабее другой.
Половая система обнаруживает у брюхоногих большие вариации (переднежа-берные обычно раздельнополы, легочные и заднежа-берные — гермафродиты). Низшие брюхоногие не имеют специальных половых протоков, и половая железа открывается у них в правую почку.
Половая железа всегда одна. У раздельнополых форм (рис. 453) это яичник или семенник, у гермафродитов — гермафродитная железа, в которой образуются и живчики и яйца. Выводные протоки Prosobranchia сравнительно просты. Самец имеет семяпровод, открывающийся наружу близ переднего конца тела на правой стороне. Либо у самого отверстия, либо впереди него на голове помещается мускулистый вырост — совокупительный орган. У самки яйцевод может образовывать местное расширение — матку, а также семяприемник.
Половые пути Pulmonata и Opisthobranchia несравненно сложнее, как можно видеть на примере виноградной улитки (см. рис. 446). От гермафродитной железы отходит общий гер-мафродитный проток, принимающий в себя выводной канал особой белковой железы. После впадения в проток железы он расширяется, причем главная часть его просвета выполняет роль яйцевода, а узкий желоб, проходящий по одной стороне яйцевода, проводит семя. Далее этот общий проток разделяется на два самостоятельных канала: яйцевод и более тонкий семяпровод. Семяпровод переходит в мускулистый совокупительный
орган (пенис). Яйцевод расширяется и образует матку, в которую впадают протоки пальцевидных желез. Матка при помощи влагалища открывается в особое впячивание стенки тела — половую клоаку, куда открывается и мешок совокупительного органа. Кроме того, во влагалище впадают еще два мешковидных образования — семяприемник, предназначенный для восприятия семени другой особи, и мешок с «любовной стрелой»; стрела — это игла из углекислой извести, которая при копуля-
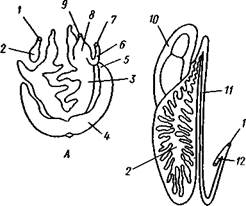
Рис. 454. Развитие морского блюдечка Patella (Prosobranchia, Diotocardia) (по Паттен). А — трохофора; Б — парусник до скручивания; В — парусник после скручивания: 1 — теменной султан, 2 — реснички верхнего полушария, 3 — прототрох, 4 — рот, 5 — просвечивающие мезодермальные полоски, 6 — раковина, 7 — внутренностный мешок, в — мантия, 9 — реснички задней части тела, 10 — зачаток ноги, 11 — кишечник ции втыкается в кожу другой особи и служит для ее раздражения. Оплодотворение перекрестное. Развитие.Среди примитивных брюхоногих встречаются формы, обладающие наружным оплодотворением, у большинства же форм оплодотворение внутреннее. Яйца часто окружаются пергаментными или студенистыми коконами и соединяются в кладки. Дробление яйца чрезвычайно напоминает таковое Polychaeta (с. 261). Оно полное, неравномерное, детерминированное, протекает по спиральному типу. Имеются те же макромеры А, В, С и D с четырьмя квартетами микромеров. Главная часть мезодермы тоже формируется за счет клетки D — из ее потомка 4d. Целомическая мезодерма образует две мезодермальные полоски, происходящие от двух первичных мезодермальных клеток. У низших Prosobranchia из яйца развивается настоящая трохофора с предротовым венчиком ресниц (прототрохом) и с теменным султаном (рис. 454, Л). Затем трохофора превращается в личинку парусник, или велигер (рис. 454, В). У большинства представителей подкл. Prosobranchia и Opisthobranchia из яйца выходит сразу парусник. Парусник часто характеризуется развитием на предротовом отделе тела 2—4 боковых лопастей, окаймленных предротовым венчиком ресниц. Лопасти образуют мерцательный парус, от которого и сама личинка получила свое наименование. Парусник обнаруживает ряд признаков, специфичных для моллюсков. На его спинной стороне появляется утолщение эктодермы, которое затем углубляется и дает легкое впячивание — раковинную железу. На дне впячивания выделяется нежный роговой зачаток раковины; впячивание затем выворачивается, зачаток раковины занимает поверхностное
положение и разрастается. На брюшной стороне тела, позади рта личинки, образуется выступ, нога, на которой в виде двух маленьких боковых впячивании эктодермы закладываются статоцисты. Впереди рта появляются щупальца и глаза. Позади зачатка раковины эктодерма образует складку, нависающую над порошицей и прикрывающую зачаток мантийной полости. У легочных мантийная складка продолжает расти вперед и становится глубже лишь на правой стороне тела, постепенно перемещаясь к переднему концу. У низших брюхоногих вместо этого происходит быстрый поворот зачатка мантийной полости с порошицей и раковины на 180° (рис. 454, Б, В), вследствие чего порошица ложится на переднем конце туловища, позади головы. Свободноплавающий парусник испытывает в конце превращения редукцию предротового венчика ресниц, опускается на дно и переходит к ползающему образу жизни. Описанный метаморфоз характерен для большинства представителей подклассов Prosobranchia и Opisthobran-chia; у Pulmonata развитие прямое и протекает до конца внутри яйцевой оболочки.
Происхождение асимметрии.Ряд ученых старались разгадать происхождение асимметрии у брюхоногих, но часть высказанных гипотез ныне имеет лишь исторический интерес.
Наиболее современной и обоснованной является теория Нэфа (1913). Есть все основания предполагать, что предками класса Gastropoda были вполне симметричные моллюски с порошицей и мантийным комплексом на задней стороне тела (рис.455), но имели раковину, закрученную в одной плоскости. Такая раковина нисколько не нарушала билатеральной симметрии. Так как вход в мантийную полость, а следовательно, и устье раковины находились сзади, то завиток раковины помещался впереди, над головой. Такая раковина несвойственна современным брюхоногим, но она характерна для примитивного головоногого моллюска — кораблика (Nautilus, с. 486). Кораблик ведет плавающий образ жизни, его раковина с завитком, обращенным вперед и закрученным в одной плоскости, не препятствует плаванию. При ползании по дну такая раковина, напротив, была бы значительной помехой, завиток ее давил бы на голову, а край устья скреб бы дно. Поэтому естественно допустить, что предок брюхоногих вел плавающий образ жизни. Причиной появления характерных черт организации Gastropoda, по Нэфу, послужил переход их предка от плавающего к ползающему образу жизни. При этом сразу же проявились отмеченные неудобства рако-кины с завитком, обращенным вперед. Устранение этих неудобств происходило следующим образом. Характерной особенностью организации брюхоногих и их ближайших предков является разделение тела на 2 области — голову с ногой и внутренностный мешок, связанные узким
мускулистым стебельком — основанием внутренностного мешка. Относительное положение главных частей тела может значительно изменяться благодаря мышечным сокращениям стебелька. Это прекрасно видно на современных брюхоногих, способных поворачивать раковину с лежащим внутри нее внутренностным мешком. При этом голова и тесно связанная с ней нога сохраняют свое нормальное положение, а внутренностный мешок может мышечными усилиями поворачиваться вокруг вертикальной оси на 180° и более. Такие временные повороты внутренностного мешка на стебельке называются физиологической тор-сией (скручиванием).
Перейдя к ползанию по дну, предок Gastropoda вынужден был повертывать внутренностный мешок с раковиной (рис. 456). Поворот на 180° оказался наиболее выгодным. Сначала он носил характер временной физиологической торсии, но потом новое положение раковины — завитком назад, а устьем вперед — стало постоянным и закрепилось наследственно. Именно такое положение раковины и характерно для современных представителей кл. Gastropoda. Замечательно, что молодой парусник низших переднежаберных имеет сначала раковину, обращенную завитком вперед (см. рис. 454,5), мантийные органы развиваются у него на задней стороне тела. Затем, опустившись на дно и переходя к ползанию, он испытывает торсионный процесс — вследствие сокращения мышц раковина с внутренностным мешком сразу (иногда в течение нескольких минут) поворачивается на 180° вершиной назад (см. рис. 454,В). У высших форм класса Prosobran-chia этот процесс происходит на более ранних стадиях развития и сводится к неравномерному росту: левая сторона основания внутренност-
Рис. 457. Раковина Tremanolus (Bellerophontidae) (из Мура). А — снизу; сверху; В — сбоку
ного мешка растет быстрее правой. Поэтому у них из яйца выходит парусник с раковиной, обращенной вершиной назад и с передним положением мантийного комплекса. Итак, у первичных древних брюхоногих раковина повернулась на 180°. Мантийные органы при этом оказались над головой на передней стороне туловища, а длинные плсвровисцеральные коннективы неизбеж-
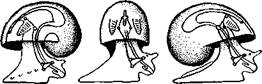
Рис. 458. Схема перехода к асимметричной па-ковине. Вид спереди (по Нэфу). А и Б — последовательные стадии: / — ктенидии, 2 — желудочек сердца, 3 — предсердия. 4 — задняя кишка Рис. 459. Схема регуляции положения раковины. Вид спереди (по Нэфу). А — первичное состояние, ось раковины поперечна; Б — ось раковины занимает косое положение, вершина сдвинута назад; В — нормальное положение раковины, вершина приподнята кверху но перекрестились, т. е. возникла хиастоиеврия (рис. 456). Раковина, повернувшаяся вершиной назад, сперва оставалась вполне симметричной. Именно такими брюхоногими, по-видимому, были ископаемые Bel-lerophontidae из нижнего палеозоя (рис. 457). Следовательно, раковина, повернутая вершиной назад, переднее положение мантийного комплекса и хиастоневрия возникли одновременно как результат единого процесса филогенетической торсии.
Дальнейшие эволюционные изменения в пределах класса Gastropoda сводятся к следующему. В результате ползающего образа жизни увеличилась нога и соответственно должен был возрасти объем раковины, в которую при опасности втягиваются как голова, так и нога. Увеличение объема раковины при сохранении прежнего ее диаметра возможно лишь за счет расположения ее оборотов в разных плоскостях. Поэтому раковина приняла коническую форму, а вершина ее все более и более стала выдаваться вправо (рис. 458). Но такая асимметричная (правозакручен-ная) раковина имеет неустойчивое положение, перетягивая животное на правый бок. По-этому для сохранения равновесия онг все более отклоняется вершиной назад и вверх, а устье ее повертывается несколько вправо (рис. 459), г. е. происходит некоторое раскручивание, или деторсия. Это яв- „ ... „ Рис. 460. Схема образования асимметрии ман- ление, называемое регуляцией ТИЙНОГо комплекса. Вид спереди (по Нэфу). пприл;М7Тттрртприил i-n / —ктенидии, 2 — желудочек сердца, 3 — левое пред-
правую сторону мантийной полости. 2. Недостаток места в правой части мантийной полости и повышенное давление вызывают постепенную редукцию и, наконец, полную утрату правых мантийных органов (рис. 460). Так возникают характерные черты Prosobranchia Monoto-cardia (рис. 461), которые лишены правых ктенидиев, осфрадия, предсердия и почки.
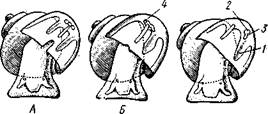
Рис. 461. Схема типов асимметрии у разных Gastropoda (по Штемпелю). А — Prosobranchia Diotocardia; Б — Prosobranchia Monotocardia; В — Pulmonata; Г — Opisthobranchia: / — рот, 2 — церебральный ганглий, 3—плевральный ганглий, 4—париетальный ганглий, 5 — висцеральный ганглий, 6. 7 — осфрадии, 8, 9 — ктеиидии, 10 — печень, // — половая железа. 12 — анус, 13 — перикардий, 14 — сосудистая Сеть легкого При сильной деторсии (у подкл. Opisthobranchia; рис. 461.Г) мантийный комплекс оказывается на правой стороне тела, а нервная система раскручивается, вновь становясь эвтиневральной. У легочных (подкл. Pulmonata; рис. 461,5) деторсия незначительна, и мантийные органы остаются в передней части тела, их нервная система также эвтиневраль-на, но эта особенность вызвана не деторсией, а сильной концентрацией ганглиев в области глотки. Позднее Гарстранг (1928) внес существенное исправление в теорию Нэфа. По его мнению, торсионный поворот внутренностного мешка у предков Gastropoda происходил не во взрослом состоянии, а еще у планктонных личинок, как это наблюдается у низших современных брюхоногих (например, у Patella; рис. 462,Г). Этот процесс был полезен, так как только обращенная устьем вперед раковина давала возможность личинке втягивать в нее сначала более жизненно важную часть тела — голову, а потом уже ногу. Итак, это исправление Гарстранга касается только самого первого этапа становления асимметрии, все остальные и более поздние ее проявления, по-видимому, были правильно объяснены Нэфом. Экология.Большинство Prosobranchia и все Opisthobranchia — морские животные. Некоторые переднежаберные приспособились к жизни в пресной воде и даже на суше (Viviparus, Bithynia, Cyclophorus, Acme, Helicina и др.)- В подклассе Pulmonata представлены в основном сухо-
путные и пресноводные формы. В море брюхоногие встречаются на различных глубинах, на суше — в самых различных климатах — выносливы по отношению к температуре. Перенесение резких колебаний температур облегчается способностью многих легочных к спячке — зимней на севере, летней и зимней на юге. При этом улитка забирается в почву, втягивается в раковину и заклеивает устье эпифрагмой, т. е. пленкой из застывшего слоя слизи с большим содержанием солей извести. Переход от ползания по дну водоема к плаванию совершался у брюхоногих в различных группах независимо. Пелагическими брюхоногими являются киленогие (см. рис. 439) —Heteropoda (из подкл. Prosobranchia) и крылоногие (см. рис. 440)—Pteropoda (из подкл. Opisthobranchia); и те и другие хорошо приспособлены к плаванию в толще воды. Тело их обычно стекловидно, прозрачно, нога образует непарный килевидный плавник (Heteropoda) или превращена в пару больших крыловидных плавников (Pteropoda); раковина более или менее редуцируется, вплоть до полного исчезновения. Настоящие паразиты имеются только среди переднежаберных брюхоногих. Большинство их паразитирует на коже или в полости тела иглокожих (морские звезды, морские ежи, голотурии). Паразитический об раз жизни вызвал у этих брюхоногих (Entocolax, Parenteroxenos) сильные изменения и упрощения организации (утрата раковины, мантии, ноги и т. д. вплоть до полной редукции пищеварительной, кровеносной, нервной систем). Пища брюхоногих разнообразна. Наряду с растительноядными формами, широко распространенными в воде и на суше, имеется большое количество хищников, питающихся червями, раками или другими моллюсками. Полезное и вредное значение брюхоногих.Практическое значение брюхоногих по сравнению с другими моллюсками (пластинчатожаберными и особенно головоногими) не очень велико. В ряде европейских стран употребляется в пищу виноградная улитка (Helix pomatia), которую для этой цели разводят в специальных улитковых хозяйствах. Съедобны и многие морские переднежаберные: трубач Buccinum undaturn, береговая улитка Littorina и некоторые другие. Перламутр раковины некоторых морских Prosobranchia, например Turbo, Trochus и Haliolis, служит для выделки различных украшений, безделушек и пуговиц. Различные морские раковины, главным образом каури (Monetaria moneta), вплоть до начала XX в. служили у некоторых народов в качестве разменной монеты. Сведения о подобном использовании каури имеются, например, в летописях Индии от VII в. н. э. В XIX в. каури ввозились в большом количестве в Западную Африку для сбыта туземцам, делавшим из нанизанных в, виде бус раковин денежную единицу. Вредное значение для сельского хозяйства имеют некоторые наземные формы из подкласса Pulmonata, а именно слизни, или слизняки. Примером слизней может служить достигающий 3—6 см длины полевой слизень (Deroceras reticulatus), наносящий вред озимым посевам, а также картофелю, свекле, табаку, клеверу и огородным растениям. В южных районах значительный вред могут наносить моллюски рода Parmacella, относящиеся к группе слизней. Многие из них представляют серьезную угрозу для садов и огородов. Велико отрицательное значение брюхоногих как первых промежуточных хозяев трематод (с. 168), среди которых имеется много возбудителей опасных заболеваний человека и животных. Палеонтология.Наиболее древние брюхоногие, известные с кембрия, - относятся к переднежаберным. Среди них интересны еще сохранившие симметричную раковину Bellerophontidae (ордовик — пермь) и дожившая с триаса до наших дней Pleurotomaria (рис. 462,Л). В карбоне появляются заднежаберные и легочные моллюски. Такие распространенные сейчас пресноводные роды, как прудовики и катушки, ведут свое начало с юры (p. Lymnaea) и мела (p. Planorbis). Классификация.Большое количество видов брюхоногих моллюсков распределяется между тремя подклассами. Классификация в пределах последних отличается сложностью в связи с большим разнообразием в строении брюхоногих.
Поиск по сайту: |