|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
КЛАСС II. НАСЕКОМЫЕ (INSECTA)
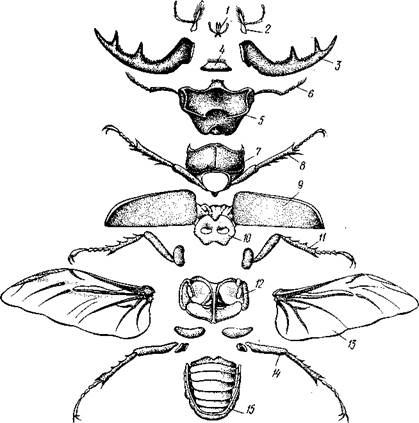
Класс насекомых включает всех трахейнодышащих членистоногих, обладающих тремя парами ног. Тело их отчетливо подразделяется на голову, грудь и брюшко. Большинству представителей класса присуща способность к полету — единственная в этом смысле группа среди всех беспозвоночных. Класс насекомых богаче всех прочих классов по числу представителей. На долю Insccta приходится около 70% общего числа известных видов животных. По разным источникам число изученных видов насекомых колеблется от 600000 до 1 500000. Тем не менее и до сих пор в научной литературе ежегодно описываются новые формы. Если принять во внимание, что многие виды насекомых встречаются в громадном количестве особей, то становится понятным, какую огромную роль насекомые играют в наземных биоценозах. Естественно, что и практическое значение насекомых чрезвычайно велико. Строение и физиология.Тело взрослых Insecta разделено на голову, грудь и брюшко (рис. 307). Сегменты головы слиты в общую массу, тогда как членики груди и брюшка бывают более или менее хорошо различимы. Голова состоит из акрона и 4 сегментов, грудь всегда из 3, брюшко же в своем наиболее полном составе содержит 11 сегментов и тельсон. Голова и грудь несут конечности, брюшко иногда сохраняет лишь их рудименты. Голова одета общей хитиновой капсулой, резкий пережим, или шейка, отделяет ее от груди, с которой она большей частью соединена под-
Рис. 307. Расчленение жука-оленя Lucanus cervus (из Гилярова): / — нижняя губа, 2 — нижняя челюсть. 3 — мандибула. 4 — верхняя губа, 5 — голова, 6 — сяжки, 7 — передпегрудь, 8 — первая пара грудных конечностей, 9 — передняя пара крыльев — надкрылья, 10 — среднегрудь, // — вторая пара грудных конечностей, 12 — заднегрудь, 13 — задние крылья, 14 — третья пара грудных конечностей, 15 — брюшко
Рис. 308. Голова насекомого спереди (Л) и сбоку (Б) (из Кенигсмена): / — головная капсула, 2 — простые глазки, 3 — фасетированный глаз, 4 — антенна (сяжки), 5 — мандибула, 6 — верхняя губа, 7 — место прикрепления сяжка, S — : нижняя губа, * — нижняя челюсть вижно. На нижней поверхности головы или на ее переднем конце помещается рот. По бокам головы расположены два больших сложных глаза, между которыми могут находиться несколько мелких одиночных глазков (рис. 308).
Голова несет 4 пары придатков. От верхней стороны ее отходит пара антенн — придатков акрона. Они гомологичны таковым многоножек, но более разнообразны по форме. Их также часто называют «усиками», или «сяжками». Сяжки всегда состоят из одного ряда члеников. Различают щетинковидпыс сяжки, нитевидные, пильчатые, гребенчатые, перистые, коленчатые и др. (рис. 309). Усики снабжены многочисленными рецепторами, воспринимающими различные типы раздражений. Главным образом это органы осязания и обоняния.
Интеркалярный сегмент (I сегмент тела) не только не обладает никакими придатками, но и подвергается почти полной редукции. Прочие три пары придатков — конечности II, III и IV сегментов, вошедших в состав головы. Конечности располагаются вокруг ротового отверстия и в совокупности с рядом других образований, имеющих независимое от конечностей происхождение, формируют довольно сложно устроенные у насекомых ротовые аппараты; последние представляют единое функциональное целое. В соответствии с крайним разнообразием способов питания ротовые аппараты многих групп насекомых отличаются по строению. Однако изучение морфологии и принципов действия этих образований показывает, что многообразные ротовые части Insecta — результат изменений одного общего основного типа. Последний, в свою очередь, возник путем специализации обычных конечностей членистоногих. Захват, удержание и измельчение пищи становятся их главной задачей. Решая вопрос об исходном у насекомых типе ротовых аппаратов, естественно исходить как из строения, так и из функциональных особенностей последних. Общность плана строения ротовых конечностей и ход-ной ноги, легкость установления гомологии между их отдельными частями— показатели примитивности ротового аппарата в целом. О многом может сказать и характер пищи. По-видимому, первичным для насекомых было питание более или менее твердым органическим детритом растительного или животного происхождения, что определило появление хищничества или настоящей растительноядности. Но во всех этих случаях животному необходимо захватывать и разгрызать пищевой объект. Поглощение жидкой пищи (кровь животных, соки и нектар растений, жидкие разлагающиеся органические вещества и т. п.) обусловило появление принципиально иных приспособлений.
Этим условиям полностью отвечают встречающиеся у прямокрылых, тараканов, жуков и т. п. ротовые аппараты грызущего типа. Их первичный характер доказывается наиболее богатой расчлененностью отдельных конечностей, сходством с ротовыми органами многоножек и тем, что они встречаются у личинок многих отрядов насекомых (например, чешуекрылых) , у которых во взрослом состоянии ротовые аппараты других типов.
В состав ротового аппарата грызущего типа (рис. 310) входят следующие образования. Спереди рот прикрыт «верхней губой», представляющей складку покровов головы и образующей переднюю стенку ротовой полости. Верхняя губа закладывается независимо от конечностей и не гомологична им. За ней следует пара верхних челюстей— жвал, или мандибул. Это две толстые нерасчлененные пластинки, зазубренные по внутреннему краю и играющие наиболее важную роль в размельчении пищи. Мандибулы являются конечностями II сегмента головы. Конечности III и IV сегментов — максиллы, или нижние челюсти. Они члени-сты н этим напоминают О происхождении ОТ ходных ног. Первая пара нижних челюстей г г раСПОЛОЖеНЗ ПО бОКЗМ рТЗ, ПрИЧбМ КЭЖДЗЯ челюсть СОСТОИТ ИЗ ДВуЧЛбНИКОВОГО ОСНОВа- ния и сидящих на его вершине трех придатков: двух лопастей, внутренней и наруж- НОЙ- И ЧЛСНИСТОГО Ч6ЛЮСТНОГО ЩупИКЗ. ЭтО НЗИбоЛСС богЗТО раСЧЛеНбННЫе КОНСЧНОСТИ насекомых, вследствие чего как раз на строении первой пары максилл базируются попытки провести гомологию между конечностями Insecta и основным двуветвистым типом конечностей другой ветви членистоногих, а именно ракообразных. Вторая пара нижних челюстей насекомых сливается (как у многоножек Chilopoda, см. рис. 302) и образует непарную пластинку — нижнюю губу. Соответственно происхождению нижняя губа состоит из основной пластинки и сидящих на ней трех пар придатков. Последние представляют собой две пары нерасчлененных лопастей и пару нижнегубных щупиков. К ротовому аппарату относится также хитиновое выпячивание дна ротовой полости — гипофаринкс, или язык. Наименьшее сравнительно с описанным основным типом изменение обнаруживают лакающие ротовые органы (рис. 311) многих перепончатокрылых (пчелы, шмели). Верхняя губа и жвалы приблизительно такого же строения, как и в грызущем ротовом аппарате. Напротив, обе пары нижних челюстей заметно изменяются по сравнению с исходным типом: сохраняя полный набор частей, входящих в их состав, они сильно вытягиваются в длину и в сложенном «рабочем» состоянии образуют довольно широкий хоботок. Подобная двойственность в строении ротового аппарата пчел и шмелей объясняется особенностями их питания. Мандибулы служат для сбора и размалывания твердой цветочной пыльцы, а хоботок, образуемый максиллами, — для всасывания нектара. Переход многих насекомых к питанию только жидкой пищей привел к возникновению более специализированных ротовых аппаратов,
приспособленных для сосания, основу которых составляет хорошо герметизированная трубка. Последняя в разных отрядах насекомых имеет различное происхождение и строение в зависимости от особенностей источника и характера пищи. У комаров (отр. Diptera), например, совокупность всех ротовых частей образует колющий хоботок (рис. 312), состоящий из желобовидного футляра, в котором заложены колющие щетинки. Желоб образован очень сильно вытянутой нижней губой, щупики которой почти полностью атрофируются. Сверху желоб прикрыт тоже вытянутой верхней губой, края которой смыкаются, образуя узкую трубку, служащую для всасывания крови. Жвалы, две пары нижних челюстей и гипофаринкс преобразованы в тонкие колющие стилеты, легко проникающие через покровы позвоночных животных.
Сосущие ротовые органы (рис. 313) встречаются у чешуекрылых и устроены в виде сосательного хоботка. Ротовой аппарат характеризуется необычайным развитием первой пары нижних челюстей и редукцией всех остальных частей. Верхняя губа едва различима, жвал нет. Нижняя губа образует небольшую нерасчлененную треугольную пластинку, снабженную трехчлениковыми щупиками. Нижняя челюсть каждой Рис. 311. Лакающие ротовые органы шмеля Bombus (по Богданову-Катькову): / — верхняя губа, 2 — верхняя челюсть, 3 — язычок — сросшиеся внутренние лопасти нижней губы, 4 — нижнегубной щупик, 5 — наружная лопасть нижней губы, 6 — нижнечелюстной щупик, 7 — подбородок, 5 — нижняя челюсть. 9 — стволик, 1П — основной членик, // — подпод-бородок Рис. 312. Колющие ротовые органы комара. А — хоботок комара в расправленном виде (по Муру); Б — положение ротовых частей во время акта сосания крови (по Веберу); В — поперечный разрез через хоботок комара (из Зеликмана): / — верхняя губа, 2 — мандибула. 3 — нижняя челюсть, 4 — гипофаринкс, 5—нижняя губа, 6 — «ижне-челюстной щупик, 7 — сяжки
стороны вытянута в очень длинный желобок, вогнутость которого обращена к медианной линии тела. Желобки обеих сторон плотно прикладываются друг к другу краями, образуя трубку. В покоящемся состоянии хоботок свернут на брюшной стороне в крутую спираль и спрятан под
Рис. 313. Сосущий ротовой аппарат бабочки. /1—голова бабочки с расправленным хоботком; Б — участок хоботка при большом увеличении (по Веберу): / — основание усиков, 2 — верхняя губа, 3 — нижнегубной щупик, 4 — нижняя губа, 5 — хоботок — нижние челюсти, 6 — фасетированный глаз, 7 — правая и левая нижние челюсти, S — места соединения челюстей, 9 — полость хоботка, 10 — трахеи, 11 — мускулатура хоботка головой. В расправленном виде хоботок засовывается внутрь цветков при высасывании нектара. Лижущим ротовым аппаратом (рис. 314) обладают многие мухи из двукрылых. Главная часть аппарата — мясистая нижняя губа (хоботок), заканчивающаяся двумя большими пластинчатыми выростами, последние снабжены сложно устроенным фильтрующим аппаратом. Мандибулы и первая пара нижних челюстей атрофированы, хотя челюстные щупики сохраняются. Верхняя губа и гипофаринкс, расположенные в желобооб-разном углублении на передней стенке нижней губы, в совокупности с ней образуют трубочку, в которую и поступает жидкая пища, слизанная и профильтрованная пластинчатыми выростами нижней губы. В некоторых случаях может происходить вторичное упрощение или даже почти полная редукция ротового аппарата. Это главным образом характерно для форм, не питающихся на взрослой стадии (поденки, оводы). Описанные крайние ступени развития ротового аппарата связаны с основным грызущим типом ротовых конечностей рядом переходов. Положение ротовых частей на головной капсуле имеет важное систематическое значение. У подавляющего большинства форм они расположены снаружи, на поверхности головы. Эти насекомые объединяются в подкласс Открыточелюстных (Ectognatha). У небольшого числа самых примитивных форм ротовые конечности погружены внутрь особой рото- вой капсулы, так что наружу в лучшем случае торчат только их кончики. Подобные насекомые составляют второй подкласс — Скрыточелюстные (Entognatha). Грудь насекомых (см. рис. 307) состоит из трех сегментов, называемых передне-, средне- и заднегрудыо. Сегменты груди несут три пары
двигательных конечностей, прикрепляющихся между стернитом и боковой пластинкой каждой стороны (рис. 315). Конечности всегда состоят из одного ряда члеников, в которых мы отличаем от основания к концу ноги: 1) тазик, или ляжку, широкий основной членик; 2) вертлуг; 3) бедро, самый толстый членик ноги; 4) голень, обычно самый длинный из члеников; з^шгт _
Рис. 314. Лижущий ротовой аппарат мухи Musca domestica. Вид спереди и снизу (из Шван-вича); 1 — нижнечелюстной щупик, 2 — верхняя губа, 3 — гипофаринкс, А — каналы фильтрующего аппарата. 5 — ротовое отверстие, 6 — лопасти нижней губы, 7 — нижняя губа Рис. 315. Схема строения грудного сегмента (по Шванвичу): / — тсргит, 2 — боковая пластинка, 3 — стернит. 4 —тазик, 5 — вертлуг. f — бедро, 7 — голень, 8 — лапка, 9 — крыло 5) лапку, состоящую из разного числа (максимально до 5) очень маленьких члеников. Последний из них несет на конце два (реже один) коготка. В связи с разными способами движения ноги испытывают модификации,иногда очень сильные (рис. 316). Ближе всего подходят к
Рис. 316. Различные типы конечностей насекомых. А—бегательная; Б — пры-гательная; В — копательная; Г — плавательная (из Бей-Биенко) описанному типу бегательные ноги, которые и наиболее распространены у Insecta. У насекомых с прыгательными ногами, например у кузнечиков, бедро и голень задней пары ног сильно вытягиваются. У роющих насекомых все ноги, а в особенности передние, играющие главную роль при копании, укорачиваются, становятся массивными и приобретают мощное вооружение из хитиновых зубцов. Плавательные конечности сплющены в виде весла и снабжены густым рядом упругих гребных волосков (жуки-плавунцы). Конечности насекомых, представляющие систему подвижно соединенных друг с другом рычагов с большим числом степеней свободы, способны к разнообразным и совершенным движениям. Характернейшей особенностью насекомых как большой систематической группы является их способность к полету. Полет осуществляется
Рис. 317. Последовательные стадии формирования крыла бабочки. А — зачаток крыла куколки; Б — крыло только что вышедшей из куколки бабочки; В — затвердевшее крыло (из Шванвича): / — гиподерма, 2 — полость зачатка крыла, 3 — трахея, 4—кутикула Рис. 318. Схема жилкования крыла насеко- ] мого (из Кенигсмена): / — костальная жилка, 2 — субкостальная жилка, 3— радиальная жилка, 4 — медиальная жилка, I 5 — кубитальная жилка, 6 — анальные жилки, 7 — югальные жилки при помощи крыльев; в большинстве случаев их две пары и расположены они на II (среднегрудь) и III (заднегрудь) грудных сегментах. Крылья представляют, по существу, мощные складки стенки тела. Хотя полностью сформированное крыло имеет вид тонкой цельной пластинки, оно тем не менее двухслойно; верхний и нижний слои разделены тончайшей щелью, являющейся продолжением полости тела (рис. 317,В). В крыло, как и во все участки тела, заходят трахейные стволы и нервы. В местах их залегания на крыльях образуются характерные трубчатые утолщения — жилки, располагающиеся строго определенным для каждого вида образом, в результате чего возникает определенный рисунок— жилкование. В настоящее время выработана единая система названий жилок, а исходный тип жилкования представляется следующим образом (рис. 318). Вдоль крыла проходят так называемые продольные жилки: костальная, субкостальная, радиальная, медиальная, кубитальная, анальные и югальные. Все они, кроме первой, могут образовывать дополнительные ветви. Наряду с продольными имеются и поперечные жилки. Особенности строения крыльев становятся хорошо понятными при знакомстве с процессом их развития в онтогенезе насекомых (см. рис. 317). Крылья закладываются в виде мешкообразных выпячиваний кожи, в которые продолжаются полость тела и трахеи. Выпячивания сплющиваются дорзовентрально; гемолимфа из них оттекает внутрь тела, верх-
Рис. 319. Схема работы крыльев насекомого. А —стадия поднятого крыла; Б — стадия опущенного крыла (по Шванвичу): I — тергит, 2 — крыло, 3 — столбик, 4 — боковая пластинка, 5 — дорзо-вентральная мышца, 6 — продольная мышца, 7 — Основание конечности ний и нижний листки пластинки сближаются, мягкие ткани частично дегенерируют, и крыло приобретает вид тонкой перепонки. Движение крыльев у насекомых — результат работы сложного механизма и определяется, с однойстороны, особенностью сочленения крыла с туловищем, а с другой — действием особых крыловых мышц. В общих чертах основной механизм движения крыльев представляется следующим образом (рис. 319). Само крыло — это двуплечий рычаг с неодинаковой длиной плечей. С тергитом и боковой пластинкой крыло соединяется тонкими и гибкими мембранами. Чуть отступя от места этого соединения, крыло опирается на небольшой, имеющий вид столбика вырост боковой пластинки, который и является точкой опоры крылового рычага. Расположенные в грудных сегментах мощные продольные и дорзо-вентральные мышцы могут опускать или приподнимать тергит. При опускании последний давит на короткое плечо крыла и влечет его за собой вниз. В результате длинное плечо, т. е. вся несущая плоскость крыла, движется вверх. Подъем тергита приводит к опусканию крыловой пластинки (рис. 319). Небольшие мышцы, прикрепленные непосредственно к крылу, способны поворачивать его вдоль продольной оси, при этом изменяется угол атаки. Во время полета свободный конец крыла движется по довольно сложной траектории (рис. 320). При опускании крыловая пластинка расположена горизонтально и движется вниз и вперед: возникает подъемная сила, удерживающая насекомое в воздухе. При движении вверх и назад крыло располагается вертикально, что создает про-пеллирующий эффект. Количество ударов крыла в 1 с сильно варьирует у разных насекомых: от 5—10 (у крупных дневных бабочек) до 500—600 (многие комары); у
очень мелких комаров-мокрецов эта цифра достигает 1000 колебаний в 1 с. У различных представителей насекомых передние и задние крылья могут быть развиты в разной степени. Только у более примитивных насекомых (стрекозы) обе пары крыльев развиты более или менее одинаково, хотя и разнятся по форме. У жуков (отр. Жесткокрылые — Coleo-ptera) передние крылья изменяются в толстые и твердые надкрылья — элитры, которые почти не участвуют в полете и в основном служат для защиты спинной стороны тела. Настоящими же крыльями являются лишь задние крылья, которые в покоящемся состоянии спрятаны под надкрыльями. У представителей отряда клопов твердеет лишь основная половина передней пары крыльев, вследствие чего эту группу насекомых нередко называют отрядом Полужесткокрылых. У некоторых насекомых, а именно у целого отряда двукрылых, развита лишь передняя пара крыльев, тогда как от задней остаются лишь рудименты в виде так называемых жужжалец.
Среди насекомых имеются и бескрылые, которые делятся на две различные группы. У одних отсутствие крыльев есть черта, присущая им искони и указывающая на примитивность их организации. Это все представители подкласса Entognatha и наиболее примитивные из открыточелюст-ных — отряд Thysanura (см. рис. 366). Среди остальных Ectognatha тоже встречаются формы, лишенные крыльев, — вши, блохи, некоторые мухи и т. п. Однако бескрылость в этом случае — свойство вторичное: это формы, утратившие крылья вследствие паразитического образа жизни или каких-либо иных причин.
Вопрос о происхождении крыльев еще не вполне разрешен. В настоящее время одной из наиболее обоснованных представляется «паранотальная» гипотеза, согласно которой крылья возникли из простых неподвижных боковых выростов кожи — паранотумов. Такие выросты встречаются у многих члени-
стоногих (трилобиты, ракообразные), у многих ископаемых насекомых (см. рис. 362) и у некоторых современных форм (личинки термитов, некоторые богомолы, тараканы и т. д.; рис. 321). Переходом от ползающего к летающему послужил, быть может, лазающий образ жизни на деревьях, при котором насекомые, вероятно, часто делали прыжки с ветки на ветку, что способствовало дальнейшему развитию боковых выростов груди, которые вначале выполняли функции несущих плоскостей при парашютировании или планирующем полете. Дальнейшая дифференцировка и отчленение выростов от самого тела привели к выработке настоящих крыльев, обеспечивающих активный пропеллирующий полет. Брюшко — последний отдел тела насекомых (см. рис. 307). Количество сегментов, входящих в его состав, варьирует у разных представителей класса. Здесь, как и в других группах членистоногих, выявляется четкая закономерность: чем ниже в эволюционном отношении стоят те или иные представители, тем более полным набором сегментов они обладают. И действительно, максимальное число брюшных сегментов мы находим у самых низших скрыточелюстных (отр. Protura), брюшко которых состоит из 11 сегментов и заканчивается отчетливо различимым тельсоном (см. рис. 363, А, Б). У всех остальных насекомых часть сегментов редуцируется (обычно один или несколько последних, а иногда и самый первый), так что общее число сегментов может сокращаться до 10, а у высших форм (некоторые перепончатокрылые и двукрылые) до 4-5.
Брюшко обычно лишено конечностей. Однако вследствие происхождения насекомых от форм, обладавших ножками на протяжении всего гомономно расчлененного тела, нередко сохраняются на брюшке рудименты конечностей или же конечности, изменившие свою первоначальную функцию. Так, у отряда Protura, низших представителей бескрылых насекомых, имеются маленькие конечности на трех передних члениках брюшка. Сохраняются рудименты брюшных конечностей и у открыто-челюстных. У тизанур на всех сегментах брюшка имеются особые придатки— грифельки, на которых, как на полозьях, при движении насекомого брюшко скользит по субстрату (см. рис. 366, А). Одна пара . грифель-
Рис. 322. Схема строения яйцеклада насекомого (из Бей-Биенко): / — половое отверстие. 2 — створки яйцеклада, 3 — цер- кн Рис. 323. Строение покровов насекомого (из Бей-Биенко) : /— наружный слой кутикулы, 2 — средний слой кутикулы, 3 —внутренний слой кутикулы, 4 — гиподерма, 5 — базальная мембрана. 6 — клетка, образующая волосок, 7 — волосок
ков на заднем конце тела сохраняется и у тараканов. Очень широко распространены, особенно у более примитивных форм (тараканы, саранчовые и т. п.), церки — парные придатки последнего сегмента брюшка, также являющиеся видоизмененными конечностями. По-видимому, сходное происхождение имеют и яйцеклады (рис. 322), встречающиеся у многих насекомых и состоящие из трех пар вытянутых створок. Покровы насекомых, как и всех остальных членистоногих, состоят из трех основных элементов — кутикулы, гиподермы и базальной мембраны (рис. 323). Кутикула выделяется клетками гиподермы, которая у скры-точелюстных насекомых часто превращается в синцитий. Кутикула насекомых трехслойна. В отличие от таковой ракообразных (с. 296) она несет наружный слой, содержащий липопротеинозые комплексы и препятствующий испарению воды из организма. Насекомые — сухопутные животные. Интересно отметить, что у водных и почвенных форм, обитающих в атмосфере, насыщенной парами воды, наружный слой или не выражен совсем, или развит очень слабо. Механическую прочность кутикуле придают белки, задубленные фенолами. Они инкрустируют средний, основной слой. На поверхности кутикулы находятся разнообразные, подвижно сочлененные с поверхностью тела выросты — тонкие волоски, чешуйки, щетинки. Каждое такое образование обыкновенно есть продукт выделения одной крупной гиподермальной клетки. Многообразие форм и функций волосков необычайно, они могут быть чувствительными, покровными, ядовитыми. Окраска насекомых в большинстве случаев зависит от присутствия в гиподерме или в кутикуле особых красящих веществ — пигментов. Металлический блеск многих насекомых относится к числу так называемых структурных окрасок и имеет другую природу. Особенности строения кутикулы обусловливают появление ряда оптических эффектов, которые основаны па сложном преломлении и отражении световых лучей. Покровы насекомых имеют разнообразного значения железы; они бывают одноклеточными и многоклеточными. Таковы вонючие железы (на груди клопов), защитные железы (у многих гусениц) и т. д. Наиболее часто встречаются линочные железы. Их секрет, выделяемый во время линьки, растворяет внутренний слой старой кутикулы, не затрагивая вновь образующихся кутикулярных слоев. Особыми восковыми железами у пчел, червецов и некоторых других насекомых выделяется воск. Мышечная система насекомых отличается большой сложностью и высокой степенью дифференциации и специализации отдельных ее элементов. Количество отдельных мышечных пучков часто достигает 1,5 — 2 тыс. Скелетные мышцы, обеспечивающие подвижность организма и отдельных его частей по отношению друг к другу, как правило, прикрепляются к внутренним поверхностям кутикулярных склеритов (тсргитам, стернитам, стенкам конечностей). По гистологическому строению почти все мышцы насекомых поперечнополосатые. Мышцы насекомых (в первую очередь это относится к крыловым мышцам высших групп 'насекомых: перепончатокрылым, двукрылым и т. п.) способны к необычайной частоте сокращений — до 1000 раз в секунду. Это связано с явлением умножения ответа на раздражение, когда на один нервный импульс мышца отвечает несколькими сокращениями. Богато разветвленная сеть трахейной дыхательной системы снабжает кислородом каждый мышечный пучок, что наряду с заметным повышением температуры тела насекомых при полете (за счет тепловой энергий, выделяемой работающими мышцами) обеспечивает высокую интенсивность обменных процессов, протекающих в мышечных клетках. Пищеварительная система начинается небольшой ротовой полостью, стенки которой образуются верхней губой и совокупностью ротовых конечностей. У форм, питающихся жидкой пищей, она, по существу, заменена каналами (см. рис. 312,5), образующимися в хоботке и служащими для засасывания пищи и проведения слюны — секрета специальных слюнных желез. Стенки верхней части ротовой полости и следующей за ней трубчатой глотки соединяются со стенками головной капсулы с помощью мощных мышечных пучков. Совокупность этих пучков образует своеобразный мышечный насос, обеспечивающий продвижение пищи в пищеварительныю систему.
В заднюю часть ротовой полости, как правило, около основания нижней губы (максиллы II) открываются протоки одной или нескольких (до 3) пар слюнных желез. Содержащиеся в слюне ферменты обеспечивают начальные этапы процессов пищеварения. У кровососущих насекомых (муха цеце, некоторые виды комаров и т. п.) слюна часто содержит вещества, препятствующие свертыванию крови — антикоагулянты. В некоторых случаях слюнные железы резко меняют свою функцию. У гусениц бабочек, например, они превращаются в прядильные, которые вместо слюны выделяют шелковистую нить, служащую для изготовления кокона или для других целей. Пищеварительный канал насекомых (рис. 324), начинающийся глоткой, состоит из трех отделов: передней, средней и задней кишок. Передняя кишка может быть дифференцирована на несколько отличающихся по функциям и строению частей. Глотка переходит в пищевод, имеющий вид узкой и длинной трубки. Задний конец пищевода часто расширяется в зоб, особенно развитый у насекомых, питающихся жидкой пищей. У некоторых хищных жуков, прямокрылых, тараканов и т. д. за зобом помещается еще одно небольшое расширение передней кишки — жевательный желудок. Кутикула, выстилающая всю переднюю кишку, в жевательном желудке образует многочисленные твердые выросты в виде бугров, зубцов и т. п., способствующих дополнительному перетиранию пищи. Далее следует средняя кишка, в которой происходит переваривание и всасывание пищи; она имеет вид цилиндрической трубки. При начале средней кишки в нее нередко впадает несколько слепых выпячиваний Рис- 324- Пищеварительная система черного таракана (по Веберу): кишечника, или пилорических при- ;_м'юнные ж£иы. 2ЛищеволУ_аов. ДЗТКОВ, СЛуЖаЩИХ В ОСНОВНОМ ДЛЯ 4 — пилорические придатки, 5 — средняя о кишка, 6—мальпигисвы сосуды, 7 — зад- увеЛИЧеНИЯ ВСаСЫВаЮЩеИ ПОВерХНО- НЯЯ кишка, «-прямая кишка сти кишечника. Стенки средней кишки часто образуют складки, или крипты. Обыкновенно эпителием средней кишки выделяется вокруг содержимого кишки непрерывная тонкая оболочка, так называемая пери-трофическая мембрана. В средней кишке происходит окончательное переваривание и усвоение пищевых веществ. Набор пищеварительных ферментов, характерный для того или иного вида насекомых, в первую очередь зависит от пищевого режима. Кровососы, пища которых в основном состоит из белков, обладают преимущественно протеолитическими ферментами, у бабочек, питающихся нектаром, имеются ферменты, воздействующие на сахара, и т. д. У многих растительноядных форм в кишечнике поселяются сим-биотические микроорганизмы (простейшие, бактерии и др.), обеспечивающие гидролиз клетчатки. Большую роль в процессах переваривания и всасывания пищевых веществ играет, по-видимому, перитрофическая мембрана, обеспечивающая транспорт ферментов внутрь пищевой массы, а продуктов пищеварения— в полость кишечника. Кроме того, она предохраняет эпителий средней кишки от механических повреждений. Средняя кишка переходит в эктодермальную заднюю, которая нередко отличается значительной длиной и подразделением на несколько участков. Здесь у большинства насекомых развиваются так называемые ректальные железы. О функциональных особенностях этих образований, как и всей задней кишки в целом, будет сказано ниже. Пища насекомых чрезвычайно разнообразна и охватывает почти все вещества растительного и животного происхождения. Среди насекомых имеются всеядные, например тараканы, которые питаются различными растительными и животными продуктами. Очень большое количество насекомых— чистые вегетарианцы и питаются различными частями растений от корня до плода включительно. Не менее многочисленны насекомые-хищники, поедающие других насекомых, червей, моллюсков и т. д. Наконец, имеются насекомые, питающиеся падалью и разного рода отбросами и продуктами гниения: навозом, гниющими растительными остатками и т. д. Некоторые из насекомых специализировались, казалось бы, на крайне малопитательной пище: перья, рог, воск и т. д. Органы выделения. Функции органов выделения у насекомых выполняются рядом образований. В первую очередь, это мальпигиевы сосуды, отходящие от пищевого канала на границе между средней и задней кишками (см. рис. 324). Мальпигиевы сосуды Tracheata развиваются за счет задней кишки и имеют соответственно эктодермальное происхождение. Их полость выстлана низким однослойным эпителием. Количество мальпигиевых сосудов колеблется от 2 до 200 и более или менее постоянно у представителей разных систематических групп. Иногда мальпигиевы сосуды обладают некоторой подвижностью, что обеспечивает постоянную смену омывающей их гемолимфы. Из гемолимфы продукты обмена в виде растворенных в воде солей мочевой кислоты поступают в полость сосудов, где образуется труднорастворимая мочевая кислота, кристаллы которой сразу же начинают выпадать в осадок. Освобождающаяся при этом вода всасывается стенками сосудов и поступает обратно в гемолифму. Особенно интенсивно эти процессы идут в задней кишке, куда поступают продукты обмена из мальпигиевых сосудов. Упоминавшиеся выше ректальные железы являются основным местом всасывания воды. Почти сухие кристаллы мочевой кислоты вместе с непереваренными остатками пищи выводятся наружу через анальное отверстие. Описанный выше процесс выведения продуктов обмена в виде сухих кристаллов, а не в растворе, позволяет насекомым очень экономно расходовать воду, поступающую в организм. Последнее особенно важно для видов, которые существуют в условиях сильного дефицита влаги. У форм, обитающих в среде с повышенной влажностью или поглощающих большое количество жидкой пищи (например, тли, сосущие соки растений), реабсорбция воды не наблюдается. Кроме мальпигиевых сосудов выделительную роль играет жировое тело, развитое у насекомых сильнее, чем у многоножек. Это рыхлая ткань, обильно пронизанная трахеями . Часть составляющих ее клеток выполняет экскреторную функцию — в них откладывается мочевая кислота в виде круглых конкреций. Жировое тело относится к числу «почек накопления», в которых продукты обмена постепенно накапливаются, а не выводятся во внешнюю среду. Не следует, однако, забывать, что главная функция жирового тела — отложение запасных питательных веществ. Благодаря образованию таких запасов насекомые могут подолгу голодать (например, клопы — до 6 месяцев и больше). Сложные процессы метаморфоза насекомых, особенно на стадии куколки, когда организм не питается, также осуществляются за счет энергетических веществ, накопленных в жировом теле. Говоря об органах выделения, следует упомянуть о перикардиальных клетках, или нефроцитах, которые часто наблюдаются у насекомых. Это парные более или менее метамерно расположенные по бокам сердца группы клеток, способные поглощать из полости тела введенные туда посторонние вещества, например кармин. Помимо перикардиальных клеток у некоторых низших насекомых (тизануры, прямокрылые) найдены расположенные под сердцем в виде особых скоплений амебоидных клеток фагоцитарные органы. Последние способны захватывать различные мелкие твердые частицы, например зерна туши, введенной в полость тела. Видоизмененными участками жирового тела являются органы свечения некоторых насекомых, например жуков-светлячков Lampyris (Иванов червячок; рис. 325). Эти участки жирового тела залегают под прозрачным кутикулярным покровом брюшка; свечение их зависит от присутствия в клетках особого вещества люциферина. При наличии кислорода под воздействием особого фермента люциферазы происходит
Рис. 325. Светляк, или Иванов червячок, Lampyris noctiluca. А — самец; Б личинка (из Брема)
окисление люциферина, причем химическая реакция сопровождается испусканием света. Процессы свечения насекомых находятся под контролем нервной системы. Нервная система насекомых (рис. 326), как и у прочих членистоногих, исходно построена по типу брюшной нервной цепочки, однако может достигать очень высокого уровня развития и специализации. Центральная нервная система включает головной мозг, подглоточ-ный ганглий и сегментарные ганглии брюшной нервной цепочки, расположенные в туловище. Головной мозг состоит из трех участков: передний— протоцеребрум, средний — дейтоцеребрум и задний — тритоце-ребрум. Протоцеребрум и дейтоцеребрум иннервируют соответственно глаза и сяжки насекомых, т. е. придатки акрона. Тритоцеребрум принадлежит вставочному, или интеркалярно-
2 А .1 - му, сегменту, который отвечает сегмен Особенно важными ассоциативными центрами головного мозга считаются «грибовидные тела», располагающиеся в протоцеребруме. Впрочем, сложность структуры характеризует мозг не всех насекомых, а главным образом тех, жизнь которых отличается сложностью и разнообразием жизненных функций. Поэтому мозг развит сложнее всего у общественных насекомых: муравьев, пчел, термитов. Эта закономерность у них прослеживается даже в пределах одного вида, представленного несколькими «кастами», отличающимися по сложности жизненных отправлений. У рабочих муравьев, например, грибовидные тела развиты значительно сильнее, чем у цариц и самцов (рис. 327).
Брюшная нервная цепочка состоит из сложного подглоточного ганглия, посылающего нервы к трем парам ротовых конечностей, из трех крупных обособленных грудных ганглиев и брюшных ганглиев, количество которых может варьировать. Наиболее полное их число — 11 — наблюдается только на самых ранних стадиях эмбриогенеза некоторых насекомых — тараканы, медведки, жуки и т. д. У большинства насекомых ганглии брюшной цепочки концентрируются в продольном направлении (рис. 328), так что во взрослом состоянии даже у
самых примитивных форм не встречается более 8 брюшных ганглиев. Причем последний, -VIII ганглий сохраняет следы своего сложного происхождения за счет слияния нескольких ганглиев. Однако у многих насекомых процесс концентрации ганглиев заходит значительно дальше (рис. 328). Возникают сложные брюшные и грудные ганглиозные массы. В ряде случаев все ганглии грудного и брюшного отделов могут сливаться, образуя массу, расположенную в груди, тогда как в брюш-
Рис. 327. Головной мозг муравья Lasius brevicornis. A — мозг рабочей особи; Б — мозг самки; В — мозг самца (по Уилеру): / — срединный лобный глазок, 2 — грибовидные тела мозга, наиболее развиты у рабочих пчел И рудиментарны у самцов, 3 — зрительные доли, 4 — зрительный нерв, 5 — обонятельные доли, 6 — сяжковый нерв. Все рисунки сделаны при одинаковом увеличении
•Б В Рис. 328. Нервная система насекомых. А — жука Lygistopterus san-guineus; Б — жука Gyrinus notator; В — мухи Sarcophaga carnaria (по Брандту)
ке остаются только нервы. Обычно нервная цепочка личинок богаче расчленена, чем таковая взрослых насекомых: взрослая пчела имеет всего 6 ганглиев вместо личиночных 10 (рис. 329). Насекомые обладают системой отходящих от головного мозга симпатических нервов, которые регулируют работу внутренних органов и мышечной системы. Практически во всех отделах центральной нервной системы (головной мозг, подглоточный ганглий, брюшная нервная цепочка) имеются нейросекреторные клетки. Синтезируемый в них нейросекрет транспортируется по аксонам в особые образования — прилежащие и карди-альные тела, а затем поступает в гемолимфу. Кардиальные и прилежащие тела располагаются над кишечником сразу же за головным мозгом. Они выполняют функции желез внутренней секреции.
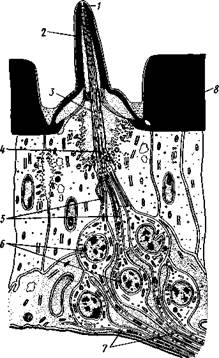
Нейросекреты играют важную роль в гормональной системе насекомых: они регулируют деятельность всех остальных эндокринных органов, гормоны которых обеспечивают нормальное осуществление развития организма, течение обменных процессов, линьки и т. д. (с. 374). Органы чувств насекомых достигают большой сложности и многообразия. Это в первую очередь определяется как общим высоким уровнем организации насекомых, так и их сложным поведением, для осуществления которого организм должен получать исчерпывающую и точную информацию об окружающем мире. Насекомые способны воспринимать самые различные раздражения — механические, звуковые, химические, зрительные и т. д. Для этого у них имеются специализированные рецепторы. Морфологической и функциональной основой органов чувств являются сенсиллы, разбросанные по телу насекомых поодиночке или собранные в более или менее крупные скопления (органы слуха, глаза и т. п.). Все сенсиллы возникли из одного исходного типа путем специализации для восприятия того или иного раздражения. Каждая сенсилла состоит из нескольких элементов. Это одна или несколько чувствительных (рецепторных) клеток. От каждой клетки берет начало центральный отросток, идущий к нервной системе, и периферический отросток. На конце последнего в большинстве случаев располагается видоизмененный жгутик, у которого отсутствуют две центральные фибриллы. Наружные части сенсиллы представлены кутикулярным образованием, строение которого зависит от выполняемых им функций. Жгутик чувствительной клетки направлен к кутикулярной части сенсиллы. Здесь его конец входит внутрь особого канала или трубочки, которые фиксируют положение жгута. Наиболее простыми являются, по-видимому, сенсиллы, воспринимающие механическое раздражение (рис. 330). Основу такой сенсиллы составляет волосок, подвижно сочлененный с кутикулой. Изменение положения волоска при соприкосновении с твердыми предметами или под
давлением воды или воздуха сразу же передается жгутику и чувствительной клетке, в которой возникает возбуждение. Такие осязательные или тактильные сенсиллы разбросаны по всей поверхности тела насекомого. По существу, механорецепторными являются и так называемые хор-дотональные сенсиллы, или сколпофоры(рис. 331), служащие для вос-
Рис. 330. Строение механорецеп-торной сенсиллы насекомых (рис. В. Иванова): / — чувствительный волосок, 2 — кутикула, :3 — кутикулярная трубочка, фиксирующая жгутик, 4 — видоизмененный жгутик, 5—периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки Рис. 331. Схема строения хордо-тоналыгой сенсиллы насекомых (по В. Иванову): / — шапочковая клетка, 2 — кутикулярная трубочка, фиксирующая жгутик, 3 —сколопоидное тельце, 4 — видоизмененный жгутик, 5 — периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки приятия колебаний различной частоты. Они в первую очередь входят в состав органов слуха насекомых. Хордотональные сенсиллы отличаются некоторыми особенностями строения. Вокруг периферического отростка чувствительной клетки и жгутика, заключенного в кутикулярную трубочку, формируется плотный фибриллярный чехол — сколопоидное тельце, имеющее вид блестящего штифтика. Сверху сенсиллу прикрывает особая шапочковая клетка, от нижней кутикулизированной поверхности которой и берет начало трубочка, фиксирующая жгут. Хордотональные сенсиллы располагаются поодиночке или группами, образуя хордото-нальные органы. Чаще всего они примыкают к внутренней поверхности мягких участков кутикулы (в сочленениях конечностей или же между сегментами тела). Иногда они погружены в глубь тела и расположены 35Z -на тонких мембранных трубках, натянутых между двумя отдельными участками кутикулы (рис. 332).
Функции хордотональных органов, по-видимому, различны. В тех случаях, когда сенсиллы примыкают к кутикуле, они, как правило, служат для восприятия низкочастотных вибраций. Правда, в отдельных случаях (хордотональные органы, расположенные в антеннах комаров) они чувствительны и к колебаниям высокой частоты. Внутренние хордотональные органы, вероятно, регистрируют изменения давления и механических напряжений, возникающих в теле насекомого. Настоящими слуховыми органами насекомых являются тимпанальные органы, в которых сколпо-форы связаны с тонкими кутикулярными мембранами (тимпанальными мембранами), играющими роль барабанных перепонок.
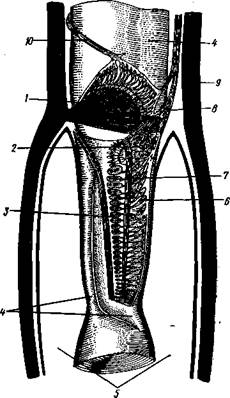
Типичным строением обладают тимпанальные органы кузнечиков (рис. 333, А, Б; 334), расположенные на голенях передних ног. В верхней части голени имеются по две узкие продольные щели, ведущие в два барабанных кармана. Внутренние стенки кармашков (рис. 333,£), обращенные друг к другу, тонки и представляют собой барабанные перепонки, наружные же утолщены и называются барабанными крышечками. Между обеими барабанными перепонками, вплотную примыкая к ним, проходят два трахейных ствола, которые, быть может, служат в качестве резонаторов. Наконец, главную часть тимпанального органа составляют три группы сколпофоров (рис. 334). Сколпофоры примыкают частью к барабанной перепонке, частью к резонирующей трахее. Центральные отростки чувстви-Рис 332 Внутренний тельных клеток образуют тимпанальный нерв. Точно по такому же принципу — сочетание сколпофоров и тимпанальных перепонок — устроены тимпанальные органы и других насекомых — саранчовых, сверчков, бабочек и др. Правда, располагаться они могут в разных местах тела — на передних сегментах брюшка, у основания крыльев и т. п. Хордотональные сенсиллы тимпанальных органов служат для восприятия колебаний различной частоты — имеются «высокочастотные» и «низкочастотные» сенсиллы. Как правило, одна из таких групп настроена на частоты, максимально представленные в звуках, издаваемых особями того же вида. В целом насекомые воспринимают звуки в очень широком диапазоне: от инфразвука (8—10 Гц) до ультразвука (45000 Гц). Насекомые способны не только воспринимать, но и издавать звуки. Эта особенность характерна для представителей многих групп: прямокрылых, жуков, перепончатокрылых, бабочек и др. Звуковые органы насекомых очень разнообразны. Стрекотание прямокрылых, например, вызывается развитием известных стрекочущих приспособлений, которые чаще всего связаны с крыльями. Так, у кузнечиковых эти органы находятся на передних крыльях. Некоторые жилки левого крыла становятся зазубренными и превращаются в так называемый смычок, которым животное водит по правому кры- лу, где в соответствующем месте находится резонатор. Последний состо ит из ограниченной высокой жилкой площадки на крыле — зеркальца. Движение зазубренного смычка по краю зеркальца приводит к вибрации растянутой на нем части поверхности крыла. У саранчовых смычок образован рядом мельчайших зубчиков на бедрах задних ног. При трении бедер о верхние крылья зубчики задевают за сильно выдающуюся у самца радиальную жилку крыла. У самцов цикад есть своеобразный «голосовой аппарат» на нижней стороне заднегруди: действие его основано на чрезвычайно быстром колебании хитиновой перепонки, приводимой в движение сокращением мышц. Значение способности издавать звуки заключается, по-видимому, в привлечении стрекочущими самцами самок.
Хеморецепторы насекомых представлены обонятельными и вкусовыми сенсиллами. Кутикулярные образования обонятельных сенсилл (рис. 335, А, Б) очень разнообразны по форме: щетинки, конусовидные придатки, пластинки и т. п. Общая черта — наличие тонких пор, пронизывающих кутикулу. Через эти поры открыт доступ к чувствительным элементам сенсиллы для молекул пахучих веществ. Обонятельные сенсиллы располагаются главным образом на сяжках и челюстных щупиках. Обоняние служит насекомым как для отыскания пищи, так и при спаривании: самцы часто находят самок по запаху. Последние выделяют особые пахучие вещества — половые аттрактанты. Достаточно ничтожного количества (100 моле-
Рис. 333. Тимпанальный орган кузнечика. А — передняя голень спереди, видны два отверстия тимпанального органа (2); Б — поперечный разрез через голень в области тимпанального органа (по Швабе): / — кутикула голени, 2 — слуховая щель, 3 — шапочковая клетка, 4 — сколпофор, 5—барабанная перепонка, в — трахеи, 7— барабанная полость, 8 — полость ноги, 9 — сколопоид-ное тельце Рис. 334. Тимпанальный орган кузнечика, вскрытый с передней стороны (по Швабе): 1 — первая группа сколпофоров (подколенный орган), 2 — вторая группа сколпофоров (промежуточный орган). 3 — третья группа сколпофоров (слуховой гребень), 4 — трахейные стволы, 5 — барабанные полости, 5 — чувствительные клетки сколпофоров, 7 — сколопоид-ное тельце, * — тимпанальный нерв, 9 — кутикула, 10 — подколенный нерв
кул в 1 см3 воздуха) такого вещества, чтобы вызвать возбуждение у самцов шелкопряда. Вкусовые сенсиллы располагаются у насекомых на ротовых конечностях и дистальных члениках лапок. Их кутикулярные элементы представлены волосками или конусовидными придатками и также пронизаны порами. В состав каждой сенсиллы (рис. 336) входит несколько рецеп-
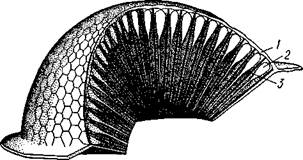
Рис. 335. Схема строения обонятельной сенсиллы насекомых. А — строение обонятельной сенсиллы (по В. Иванову); Б — различные типы обонятельных сенсилл (по Эрнсту): ; — кутикулярная дисковидная шапочка-пластинка, 2 — поры. 3 — кутикулярная трубочка, фиксирующая жгутик, 4 — видоизмененный жгутик, 5 — периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки, 8—кутикула торных клеток, каждая из которых реагирует на определенный вкусовой раздражитель: одна клетка реагирует на соли, другая на сахаристые вещества, третья на чистую воду. Одна из чувствительных клеток вкусовой сенсиллы является механорецепторной. Таким образом, у насекомых, так же как и у позвоночных, вкусовое ощущение сопровождается осязательным. Наиболее сложными из органов чувств у насекомых являются органы зрения. Последние представлены образованиями нескольких типов, из которых важнейшие — сложные фасетированные глаза примерно такого же строения, как и сложные глаза ракообразных. Глаза состоят из отдельных омматидиев (рис. 337), количество которых определяется главным образом биологическими особенностями на- \ секомых. Активные хищники и хорошие летуны, стрекозы обладают! глазами, насчитывающими до 28 000 фасеток в каждом. В то же время \ муравьи (отр. Перепончатокрылые), особенно рабочие особи видов, обитающих под землей, имеют глаза, состоящие из 8—9 омматидиев. Каждый омматидий представляет совершенную фотооптическую сенсиллу (рис. 338, А, Б). В его состав входят оптический аппарат, включающий роговицу, — прозрачный участок кутикулы над омматидием и так называемый хрустальный конус. В совокупности они выполняют роль линзы. Воспринимающий аппарат омматидия представлен несколь-
Рис. 336. Схема строения вкусовой сенсиллы насекомых (по В. Иванову): 1 — пора, 2 — кутикулярный конус, 3 — кутику-лярная трубочка, фиксирующая жгутик, 4 — видоизмененный жгутик, 5—периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки, 8 — кутикула Рис. 338. Схема строения омма-тидиев дневных насекомых (А) и ночных и сумеречных насекомых (Б) (из Мазохина-Порш-някова): / — прозрачный участок роговицы — хрусталик, 2 — хрустальный конус, 3 — пигментные клетки, 4 — рабдом. 5 — чувствительные клетки, 6 — ба-зальная мембрана, 7 — центральные отростки чувствительных клеток
Рис. 337. Схема строения сложного глаза насекомого. Участок глаза вырезан (по Гессе): / — роговица — прозрачная кутикула. ? — xpvc-тяльный конус. 3 — пигмент между омматиди-ями
кими (4—12) рецепторными клетками; специализация их зашла очень далеко, о чем говорит полная утрата ими жгутиковых структур. Собственно чувствительные части клеток — рабдомеры — представляют скопления плотно упакованных микроворсинок, располагаются в центре омматидия и тесно прилегают друг к другу. В совокупности они образуют светочувствительный элемент глаза — рабдом. По краям омматидия залегают экранирующие пигментные клетки; последние довольно существенно отличаются у дневных и ночных насекомых. В первом случае пигмент в клетке неподвижен и постоянно разделяет соседние омматидии, не пропуская световые лучи из одного глазка в другой. Во втором случае пигмент способен перемещаться в клетках и скапливаться только в их верхней части. При этом лучи света попадают на чувствительные клетки не одного, а нескольких соседних омматидиев, что заметно (почти на два порядка) повышает общую чувствительность глаза. Естественно, что подобного рода адаптация возникла у сумеречных и ночных насекомых. От чувствительных клеток омматидия отходят нервные окончания, образующие зрительный нерв. Кроме сложных глаз многие насекомые имеют еще и простые глазки (рис. 339), строение которых не соответствует строению одного омматидия. Светопреломляющий аппарат линзообразной формы, сразу же под ним расположен слой чувствительных клеток. Весь глазок одет чехлом из пигментных клеток. Оптические свойства простых глазков таковы, что воспринимать изображения предметов они не могут. Личинки насекомых в большинстве случаев обладают только простыми глазками, отличающимися, однако, по строению от простых глазков взрослых стадий. Никакой преемственности между глазками взрослых особей и личинок не существует. Во время метаморфоза (с. 373) глаза личинок полностью резорбируются. Зрительные способности насекомых совершенны. Однако структурные особенности сложного глаза предопределяют особый физиологический механизм зрения. Животные, имеющие сложные глаза, обладают «мозаичным» зрением. Малые размеры омматидиев и их обособленность друг от друга приводят к тому, что каждая группа чувствительных клеток воспринимает лишь небольшой и сравнительно узкий пучок лучей. Лучи, падающие под значительным углом, поглощаются экранирующими пигментными клетками и не достигают светочувствительных элементов омматидиев. Таким образом, схематично каждый омматидии получает изображение только одной небольшой точки объекта, находящегося в поле зрения всего глаза. Вследствие этого изображение складывается из стольких световых точек, отвечающих различным частям объекта, на сколько фасеток падают перпендикулярно лучи от объекта. Общая картина комбинируется как бы из множества мелких частичных изображений путем приложения их одного к другому. Восприятие.цвета насекомыми также отличается известным своеобразием. Представители высших групп Insecta имеют цветовое зрение, основанное на восприятии трех основных цветов, смешение которых и дает все красочное многообразие окружающего нас мира. Однако у насекомых по сравнению с человеком наблюдается сильный сдвиг в коротковолновую часть спектра: они воспринимают зелено-желтые, синие и ультрафиолетовые лучи. Последние для нас невидимы. Следовательно, цветовое восприятие мира насекомыми резко отличается от нашего. Функции простых глазков взрослых насекомых требуют еще серьезного изучения. По-видимому, они в какой-то мере «дополняют» сложные глаза, влияя на активность поведения насекомых в разных условиях ос- вещенности. Кроме того, было показано, что простые глазки наряду со сложными глазами способны воспринимать поляризованный свет. Помимо пере
Поиск по сайту: |