|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
КЛАСС I. МНОГОЩЕТИНКОВЫЕ (POLYCHAETA)
Многошетинковые — самый богатый представителями класс кольчецов (около 5300 видов), которые за единичными исключениями живут в морях. Многие из них ведут активный образ жизни, ползая по дну, роясь в грунте или плавая в толще воды; другие — сидячие животные, живут в защитных трубках. Немногие представители ведут паразитический образ жизни.
ж Рис. 211. Различные многощетинковые черви. А — Nereis (ползающая форма); Б — Le-pidonotus (ползающая форма), видны чешуевидные элитры — видоизмененные спинные усики паранодий и глотка, выдвинутая из ротового отверстия; В — Phyllodoce (ползающая форма); /" — пескожил Arenicola (роющая форма), в средней части тела видны кустистые жабры, на переднем конце выпяченная глотка; Д — Spirorbis (мелкая сидячая форма, обитающая в известковой спиральной трубочке); Е — Telepus (сидячая форма, обитающая в органической трубке, укрепленной мелкими камешками), из трубки высунут передний конец животного, снабженный длинными щупальцами; Ж — Serpula (сидячая форма, живущая в известковой трубке), на переднем конце перистые «жабры»— видоизмененные пальпы и крышечка, замыкающая вход в трубку, когда животное в нее прячется (А — из Ливанова, Б и Е — из Гаевской, В — из Матвеева, Г, Ц и Ж — по Заксу) Класс полихет характеризуется следующими признаками: чувствительные придатки головной лопасти хорошо развиты, в частности всегда имеется одна пара щупиков, или пальп, которые у сидячих полихет превращены в крону щупальцевидных придатков, часто называемых «жабрами». Каждый сегмент тела несет пару примитивных ножек — парапо-дий, снабженных щетинками.
Строение и физиология.Форма тела вытянутая, лишь слегка сплющенная в спинно-брюшном направлении или правильно цилиндрическая. Туловище состоит из различного числа (от 5 до 800) сегментов (рис. 211). По количеству сегментов различаются формы малосегментные, или олигомерные (Dinophilus, рис. 212; Myzostomum и их родичи), и формы многосегментные, или полимерные (большинство представителей Polychaeta). Передний, или предротовой, участок тела — простоми-ум и задний, или анальная лопасть, — пигидиум отличаются от сегментов туловища и являются особыми, не метамерными частями тела. Сегменты туловища в более простых случаях совершенно равноценны, или гомо-номны, имеют одинаковый вид и содержат приблизительно одинаковые органы. Такая гомономность есть признак примитивной организации и лучше всего выражена у свободноподвижных, бродячих форм. Гетеро-номность, или разноценность сегментов в разных областях тела, проявляется резче всего у сидячих полихет как следствие неодинаковых условий жизнедеятельности передней части тела, высовывающейся из трубки, и задней, всегда скрытой в глубине жилища.
Тело многощетинковых кольчецов, как правило, снабжено различными придатками, служащими отчасти для
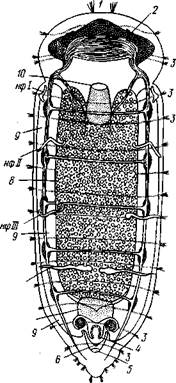
Рис. 212. Схема организации Dinophilus — олигомерного представителя Polychaeta (по Беклемишеву): / — темснняч пластинка, 2 — глаз, 3 — венчики ресничек. 4 — совокупительный орган, 5 — анус, 6—половое отверстие, 7 — задняя кишка, 8—кишечник. 9 — семенник, 10 — рот, нф! — нф!У — нефридии Рис. 213. Передний (А) и задний (Б) концы тела Nereis pelagtca (по Иванову): / — щупальце (антенна), 2 — пальп, 3 — перисто-мнальные усики, 4 — глаза, 5 — простомиум, 6 — обонятельная ямка, 7 — перистомиум, 8 — пара-подии, 9 — щетинки, 10 — спинной усик, // — пи-гидий, 12 — анальные усики, 13 — просвечивающий спинной кровеносный сосуд движения, частью органами чувств. Сильнее развиты придатки на головном отделе, где они имеют иной характер, чем на туловище. Головной отдел состоит из предротового участка — простомиума, или головной лопасти, и перистомиума, который несет ротовое отверстие и представляет первый сегмент, но часто является результатом слияния нескольких (2—3) передних сегментов (рис. 213). Процесс цефализации — включения одного или более туловищных сегментов в головной отдел — наблюдается не только у кольчецов, но и у членистоногих. Наиболее постоянные и характерные придатки простомиума — пара щупиков, или пальп. Здесь же расположена пара или более органов осязания— щупалец (антенны), имеющих разнообразную величину и форму. На перистомиуме нередко в различном количестве развиваются усики, или цирры. Пальпы и антенны иннервируются головным мозгом, усики же — передним концом брюшной нервной цепочки.
Для туловища характерно присутствие парных боковых выростов — параподий (рис. 214). Это короткие, мускулистые и подвижные выросты тела, по сути дела первые, еще очень примитивные конечности, появляющиеся у беспозвоночных. Они расположены по бокам тела метамерно, по паре на каждый сегмент. Параподия состоит из базальной нерасчлененной части и двух ветвей: спинной (нотоподия) и брюшной (невропо-дия). От основания спинной и брюшной лопастей параподий отходит по тонкому щу-пальцевидному придатку — усику, имеющему обонятельное и осязательное значение. Каждая из ветвей параподий содержит пучок щетинок, торчащих из нее концами наружу. Щетинки тонки и эластичны и состоят из органического вещества, близкого к хитину. Основаниями своими щетинки каждого пучка лежат в мешковидном впячи-вании стенки параподий. Каждая шетинка
Рис. 214. Параподия Nereis petagi- са (по Иванову): / — спинной усик, 2 — лопасти спинной ветви параподий, 3 — щетинки, 4 — лопасти брюшной ветви параподий, 5 — брюшной усик. 6 — брюшная ветвь параподий, 7 —опорные, щетинки, 8 — спинная ветвь параподий Рис. 215. Типы параподий. А — Nereis; Б — Phyllodoce; В ~ Eunice (из Абрикосова и Ле- винсона): ; — спинная ветвь, J — брюшная ветвь, 3 — спинной усик, 4 — брюшной усик, 5 — спинной усик, превратившийся в жабру, 6 — опорная щетинка
есть продукт выделения одной крупной эпителиальной клетки, залегающей на дне мешка. Нередко одна из щетинок каждого пучка отличается мощностью: это опорная щетинка; к ее основанию прикрепляются особые мышцы, приводящие в движение весь пучок. Движения параподий однообразны, они загребают спереди назад и, зацепляя щетинками за неровности субстрата, продвигают животное вперед. Форма и размеры параподий и щетинок, а также число последних сильно варьируют. Видоизменения, которым подвергаются параподий, у многих полихет выражаются большей частью в редукции спинной ветви, что ведет к образованию одноветвистых параподий (сем. Hesioni-dae, Eunicidae и др.; рис. 215). У ряда примитивных форм, например Dinophilus, выделяемых иногда в особый класс первичных кольчецов — Archiannelida, параподий и щетинки отсутствуют. Лучше всего развиты и те и другие у свободноподвижных, или бродячих, полихет. У сидячих форм в связи с их образом жизни наблюдается частичная редукция параподий; параподий нередко формируются лишь в передней части тела, высовывающейся из трубки и способной к наиболее энергичным движениям. Напротив, в задней части тела параподий как бы сглаживаются, так что щетинки торчат наружу прямо из тела. Тело Polychaeta одето однослойным кожным эпителием, который выделяет тонкую кутикулу. Эпителий местами может быть ресничным, особенно у некоторых наиболее примитивных форм. Так, у Protodrilus ресничками покрыта продольная полоса на брюшной стороне тела, а кроме того, каждый сегмент опоясан двойным венчиком ресничек: на головном участке один из венчиков лежит впереди рта, как у личинки кольчецов — трохофоры (с. 263). Часто эпителий содержит железистые клетки. У сидячих полихет выделения кожных желез могут затвердевать, образуя вокруг тела прозрачную, как- бы роговую, или хитиноидную,
Рис. 216. Схема поперечного разреза мпогощстипкового червя (из Натали): / — кожный эпителий, 2 — кольцевые мышцы, 3 — продольные мышцы, 4 — спинной усик, превратившийся в жабру, 5 — спинная ветвь параподии, 6 — опорная щетинка, 7 — мышцы параподии, 8 — воронка нефридия, 9 — канал нефридия, 10 — косая мышца, 11 — брюшной сосуд, 12 — яичник, 13 — брюшной усик параподии, 14 — брюшная ветвь параподии, /5 — кишечник, 16 — целом, 17 — спинной сосуд трубочку. Часто червь прикрепляет к органической основе трубки посторонние частицы, например, песчинки, обломки раковин моллюсков, увеличивающие ее прочность (см. рис. 211, Е). Есть, наконец, формы, у которых органическая основа трубки пропитывается углекислой известью (сем. Serpulidae; см. рис. 211, Ж). Под кожным эпителием находится хорошо развитый двухслойный мускульный мешок, состоящий из наружных кольцевых и внутренних продольных мышц (рис. 216). Продольная мускулатура у примитивных форм образует сплошной слой, у прочих же разбивается основаниями параподий на 4 продольные ленты: две из них лежат ближе к брюшной, две — ближе к спинной стороне. Внутренняя сторона продольного слоя мышц выстлана однослойным перитонеальным эпителием мезодермального происхождения, под ним находится обширная вторичная полость тела, или целом (рис. 216, 217, Б). Целом помещается между кишечником и стенкой тела, он не сплошной, а образован парными мешками, расположенными метамерно; цело-мические мешки отсутствуют в простомиуме и пигидиуме. Стенки цело-мических мешков, соприкасаясь над и под кишечником, образуют двухслойную продольную перегородку — брыжейку, или мезентерий, на которой кишечник подвешен к стенкам тела. Кроме того, на границе между
Рис. 217. Схема организации полихет. А — нервная система и нефридии с брюшной стороны; Б — кишечник и целом со спинной стороны; В — нервная система, кишечник и кровеносная система, вид сбоку (по Мейеру): / — головной мозг, 2—окологлоточный коннектив, 3—ганглий брюшной нервной лестницы, 4 — нервы сегмента, 5 — нефридий, 6 — рот, 7 — целом, 8 — кишка, 9 — диссепимент, 10 — мезентерий, 11 — пищевод, 12 — ротовая полость, 13 — глотка, /4 — мускулы — ретракторы глотки, 15 — кольцевая и продольная мускулатура, 16 —обонятельный орган, 17 — глаз, 18 — яичник, 19 и 20 — спинной и брюшной кровеносные сосуды, 21 — сплетение сосудов на кишечнике, 22 — кольцевой сосуд, 23 — сосуд глотки, 24 — пальп сегментами стенки соседних целомических мешков образуют двухслойную поперечную перегородку — септу, или диссепимент, пересекающий полость тела. Септами целом делится на известное число поперечных участков, обычно соответствующих числу сегментов тела. Впрочем, в некоторых отделах тела септы могут иногда редуцироваться. Целом заполнен водянистой жидкостью, в которой плавают амебоидные клетки. Функции целома многообразны, прежде всего он имеет опорное значение. Целомическая жидкость, по составу близкая к воде, представляет практически несжимаемое тело; при сокращении кожно-мускульного мешка стенки тела давят на целомическую жидкость, и все тело приобретает значительную упругость (ригидность). При вытягивании тела, при его сокращении, рытье норок и других типах движения значительное количество целомической жидкости перегоняется из одних отделов тела в другие (это возможно при отсутствии диссепиментов в некоторых сегментах или при наличии в них отверстий). Другие функции целома — распределительная, выделительная и половая. В целом поступают питательные вещества из кишечника, которые затем распределяются в организме. В целом поступают и в нем накапливаются жидкие и твердые экскреты — продукты обмена веществ. Наконец, в целоме происходит созревание половых клеток — спермиев и яиц. Пищеварительная система начинается ртом, который лежит на брюшной стороне перистомиума. Кишечник состоит из эктодермальной передней, энтодермальной средней и эктодермальной задней кишок. Последняя заканчивается на анальной лопасти порошицей (см. рис. 212). Передняя кишка нередко состоит из нескольких отделов. Передний из них — небольшая ротовая полость, за которой следует очень мускулистая глотка. У многих хищных видов бродячих полихет на внутренней стенке глотки кутикула местами сильно утолщается и образует острые хитиноидные зубцы, или челюстные пластинки; зубцы при выворачивании глотки служат для схватывания добычи. У нехищных сидячих форм глотка развита слабо. Средняя кишка имеет обыкновенно вид прямой трубки, задняя кишка короткая. Бродячие полихеты — преимущественно хищники и питаются различными мелкими животными; сидячие питаются главным образом взвешенными в воде органическими частицами и мелкими организмами, причем у многих для собирания пищевых частиц и для подгона их ко рту служат сильно развитые перистые головные щупики (пальпы). Они же выполняют роль жабр. Органы дыхания многощетинковых довольно разнообразны. Наиболее просто организованные представители дышат всей поверхностью тела подобно низшим червям. У большинства функцию дыхания принимают на себя известные участки параподий. Часто спинной усик превращается в жабру (см. рис. 215, В). Внутрь ее заходят кровеносные сосуды, а через стенки растворенный в воде кислород поступает в кровь. Жабры имеют листовидную, перистую или кустистую форму и развиваются большей частью не на всем протяжении тела, а только на известном его участке. Кровеносная система (см. рис. 217, В), как правило, состоит из двух главных продольных сосудов — спинного и брюшного. Один проходит вдоль всего тела над кишкой, другой — под нею, и оба залегают между двумя слоями спинного и брюшного мезентериев. Полость кровеносных сосудов представляет остатки первичной полости тела. Оба сосуда сообщаются многочисленными мелкими сосудами и лакунами, проходящими под перитонеальным эпителием кишки и кольцевыми сосудами, огибающими в стенке тела целом. Кольцевые сосуды расположены метамерно, иногда по нескольку в каждом сегменте. К ним относятся сосуды, идущие к жабрам и возвращающие кровь от жабр и от нефридиев, где кровь освобождается от продуктов распада. Главные сосуды и их ветви образуют в конце концов сеть тончайших капилляров. Последняя так густа, что, например, у пескожила (Arenicola) в ней насчитывается до 6000 ячеек на 1 мм2. Кровеносная система замкнута, т. е. капилляры, несущие кровь от главных сосудов к тканям тела, переходят непосредственно в систему капилляров, возвращающих кровь к главным сосудам. Движение крови по телу обусловливается тем, что стенки спинного сосуда, а иногда и некоторых других сокращаются и ритмически пульсируют. Общее направление тока крови в спинном сосуде идет сзади наперед, в брюшном — спереди назад. По околокишечному сплетению сосудов кровь перегоняется из спинного сосуда в брюшной, по кольцевым сосудам движется из спинного сосуда в брюшной в передней части тела и в обратном направлении в задних сегментах. Кровь нередко окрашена в красный цвет, например у пескожила (Arenicola), от присутствия в ней железосодержащего вещества, близкого к гемоглобину позвоночных. Разница лишь в том, что это вещество находится не в особых кровяных клетках, а растворено в жидкости крови. У некоторых сидячих полихет кровь зеленой окраски, зависящей от присутствия в ней хлорокруорипа, по свойствам близкого к гемоглобину. Среди многощетинковых есть небольшое количество представителей, например семейство Glyceridae, у которых кровеносная система подверглась редукции. В таких случаях функцию крови принимает на себя жидкость цслома; иногда она розового оттенка от присутствия вещества, близкого к гемоглобину. Нет кровеносной системы и у всех олигомер-ных форм (Dinophilus, Myzostomum и др.). Выделительная система полихет состоит из пефридиев. Обычно каждый сегмент туловища обладает одной парой выделительных каналов, поэтому нефридии также называют сстментарными органами (см. рис. 216; 217, Л). Внутренний конец каждого нефридия находится в целомиче-ской полости сегмента около его задней стенки, капал нефридия пронизывает диссепимент, проходит в полость следующего сегмента и затем открывается на боковой стороне тела наружу. Итак, каждый канал находится в двух сегментах, начинается в одном, по главной массой залегает в следующем.
Дальнейшие исследования показали, что нефридии многих полихет, несмотря на одинаковое положение, имеют различную природу. У ряда низших форм сегментарные органы представлены протонефридиями. В таких случаях па внутреннем конце выделительного канала сидит пучок тонких отростков, булавовидно вздутых на концах; концевой отдел канала как бы усажен пучком булавок, это особые жгутиковые клетки — соленоциты. Головка соленоцита представляет собой тело клетки с ядром; от клетки отходит длинный полый стебелек, или трубочка, открывающаяся в просвет главного канала прото-нефридия. От тела клетки в каналец торчит бьющий жгутик (рис. 218), иногда вместо одного жгутика имеется пучок жгутов. Как видно, соленоциты могут быть легко приравнены к звезд- п Л10 -,, J л. рис- 218. Конец одной из мерцательному пламени. Phyllodoce (по Гудричу) 9-763 Изучение соленоцитов с помощью электронного микроскопа показало, что они устроены совершенно сходно с терминальными клетками протонефридиев плоских червей. В обоих случаях стенка трубочки близ тела клетки имеет несколько десятков тончайших параллельных друг другу продольных щелевидных отверстий. Функция соленоцита та же, что и у звездчатой клетки протонефридиев плоских червей, т. е. осморегуляция. Выделение продуктов обмена веществ происходит через стенки главного канала протонефридия. У многих полихет вместо концевого пучка соленоцитов, который в данном случае атрофировался, появляется небольшое, усаженное ресничками отверстие, открывающееся в целом, нефридий превращается в метанефридий. Гомология метанефридиев протонефридиям подтверждается также развитием тех и других из эктодермы.
Однако дело осложняется тем, что у многих Polychaeta нефридии соединяются с половыми протоками. Исходной формой половых протоков у многощетинковых были, по-видимому, половые воронки, или цело-модукты, — короткие каналы мезодермальной природы, открывающиеся одним концом наружу, другим (воронкой) в целом. У некоторых полихет произошло прирастание половых воронок к главному каналу имевшегося в каждом сегменте протонефридия; получились смешанные нефридии, или нефромиксии (рис. 219), причем канал и соленоциты их происходят от протонефридия, воронка же — от полового протока (рис. 219,В).
Рис. 219. Нефромиксии полихет. А — Vanadis formosa, протонефри-дий и лежащая рядом независимая от него половая воронка — це-ломодукт; Б — Podarke latifrons — половая воронка, сросшаяся с каналом метанефридия; В — Alciope contrainiu — протонефридий с приросшей к нему половой воронкой (по Гудричу): I — половая воронка, 2—нефридиаль-ный канал, 3 — наружное отверстие Рис. 220. Схема взаимоотношений между нсфридиями и целомодуктами у полихет. А — гипотетическая ступень с самостоятельной половой воронкой и протонефридием; Б — нефромиксии Phyllodocidae: В — Capitel-lidae, половая воронка и метанефри-днй; Г—нефромиксии большинства полихет (по Бриану): / — целом, 2 — половая воронка. 3 — протонефридий, 4 — метанефридий
В тех случаях, когда протонефридии уже превратились в метанефри-дии, дело может обстоять двояко: иногда половые протоки и метанеф-ридии остаются обособленными; у части же полихет происходит слияние половой воронки с внутренним (отверстым) концом метанефридия. В результате получаются тоже нефромиксии, но несколько иного свойства, возникающие из слияния половых воронок не с протонефридиями, а с метанефридиями (рис. 219, Б; 220, Г). Нередко, особенно у сидячих Polychaeta, число выделительных каналов сокращается; они сохраняются всего в нескольких сегментах. Обычно, у сидячих полихет происходит физиологическая дифференциация нефромиксисв на чисто выделительные и служащие для выведения половых продуктов. Кроме нефридисв есть другие образования, играющие выделительную роль. В некоторых местах, главным образом на стенках кровеносных сосудов, перитонеальный эпителий состоит из крупных клеток, содержащих множество желтых зерен, — это накопляющиеся в клетках нерастворимые продукты обмена веществ (гуанин или соли мочевой кислоты). Наполнившись экскретами, эти, как их называют, хлорагогенные клетки отмирают, а содержимое их поступает в целом и оттуда через нефридии — наружу. Нервная система обнаруживает ряд ступеней усложнения. Типичная центральная нервная система (см. рис. 217, А) состоит из парных мозговых ганглиев, отходящих от них и огибающих глотку двух окологлоточных коннективов и парного брюшного нервного ствола. Коннективами называются нервные стволы, соединяющие разноименные ганглии (надглоточные и подглоточные или ганглии двух соседних сегментов). Поперечные нервные стволы, связывающие ганглии одного сегмента, называются комиссурами. У примитивных форм обе половины брюшного ствола еще широко расставлены и имеют равномерную обкладку из нервных клеток. Далее, у части Polychaeta правый и левый брюшные стволы начинают сближаться к срединной линии; в каждом сегменте на них появляется по скоплению ганглиозных клеток, и оба ганглия сообщаются поперечной перемычкой (комиссурой) —получается брюшная нервная система, метко названная нервной лестницей (см. рис. 212). У большинства полихет стволы сближаются еще больше, оба ганглия каждого сегмента сливаются, сами же стволы между ганглиями либо еще остаются разделенными, либо тоже сливаются, и тогда первичную парность брюшного ствола можно определить только на поперечных разрезах. Нервная система такого устройства называется брюшной нервной цепочкой. Другое постепенное усложнение нервной системы заключается в ее погружении из кожного эпителия в полость тела. Во время эмбрионального развития нервная система закладывается в виде утолщения эктодермы. У некоторых Polychaeta брюшная нервная система и во взрослом состоянии остается в кожном эпителии, не меняя своего положения. У многих полихет нервные стволы уходят под кожу и даже под кожно-мускульный мешок, располагаясь в полости тела. В некоторых случаях, когда ряд сегментов сливается друг с другом (например, в области глотки), может происходить и соответственная концентрация нервных узлов. От центральной нервной системы отходят многочисленные нервы. Головной мозг посылает от себя нервы к антеннам, пальпам и глазам, а каждый ганглий брюшной нервной цепочки иннервирует различные органы соответствующего сегмента. Органы чувств лучше всего развиты у бродячих полихет. Кроме эпителиальных чувстительных клеток, рассеянных в коже, есть специальные 9* 259 органы осязания и химического чувства. Таковыми служат антенны, пальпы, ресничные ямки, располагающиеся на простомиуме, и чувствительные усики параподий. Органы чувства равновесия редки, однако у некоторых полихет, особенно у сидячих, в передних сегментах тела имеется от 1 до 5 и более пар статоцистов. Органы зрения имеются почти у всех многощетинковых. Чаще всего глаза расположены на спинной стороне простомиума в числе 2 или 4 и
иннервируются от средней части головного мозга. Эти надмозговые глаза в наиболее простом случае представляют бокаловидное впячи-вание эктодермы с суженным отверстием. Выстилающий такую глазную ямку эпителий играет роль сетчатки, а по краям впячивания переходит в обыкновенный кожный эпителий. Сетчатка слагается из клеток двух сортов. Одни из них светочувствительны, и они-то, собственно, и являются настоящими рети-нальными клетками. Каждая такая клетка на конце, обращенном в полость впячивания, т. е. к источнику света, несет светочувствительную палочку, а на другом конце продолжается в нервное волокно. Нередко, особенно у планктонных форм, например у Alciope (рис. 221), глаз сильно усложняется: обособляется от кожи, превращаясь в замкнутый глазной пузырь, а внутри него дифференцируется, кроме двояковыпуклого хрусталика, еще прозрачное стекловидное тело. Все описанные глаза необращенного, или неинвертированного, типа. Но помимо надмозговых глаз или (при их редукции) вместо них у полихет нередко развиваются глаза на других частях тела, притом обладающие иным строением. Так, у многих сидячих форм, живущих в трубках, развиваются многочисленные глаза на пальпах, превращенных в жабры. У некоторых мелких форм (АтрЫсога и др.), ползающих задним концом вперед, развиваются глазки возле порошицы. Эти «вторичные» глаза различного строения. Половая система устроена крайне просто. Многощетинковые кольчецы раздельнополы, наружных отличий между полами нет. Половые железы формируются во всех (кроме передних и самых задних) или лишь в некоторых плодущих сегментах под слоем перитонеального эпителия. Чаще всего у основания параподий или по соседству с нефридия-ми половые клетки усиленно делятся и образуют на стенке целома местное набухание или половую железу (см. рис. 216; 217, Б). Развивающиеся гонады сначала прикрыты тонким слоем перитонеального эпителия, который впоследствии лопается, и образующиеся половые клетки попадают в целом, где они свободно плавают в полостной жидкости и достигают полной зрелости. У некоторых Polychaeta никаких половых протоков нет, так что живчики или яйца выводятся наружу просто через раз-
рыв стенки тела. У очень немногих полихет (сем. Capitellidae) имеются самостоятельные половые воронки с короткими выводными' каналами, открывающимися наружу (см. рис. 220, В). Наконец, в большинстве случаев эти воронки, как сказано, вступают в сообщение с нефридиями, и тогда нефридии несут двоякую функцию — выделение экскретов и выведение наружу половых продуктов. Оплодотворение обыкновенно наружное.
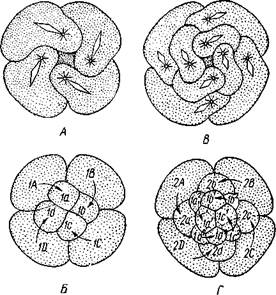
Нередко у полихет в тесной связи с половым размножением находится бесполое, приуроченное к периоду созревания половых продуктов. Переходом к этому размножению можно считать так называемые эпи-токные формы многощетинковых. Обычно наступление половой зрелости не отражается па организации червя. Иногда, однако, плодущие сегменты претерпевают сильную модификацию и в виде так называемой эпигокной части резко отличаются от остального атокного участка животного. Чаще всего эпитокной становится задняя половина червя. Изменения эпитокного участка сказываются в более сильном развитии параподий и щетинок, в сильной рудиментации кишечника, изменении окраски и т. д. Эпитокные половозрелые стадии некоторых представителей семейств Nereidae, Eunicidae и других поднимаются со дна и при помощи мощных веслообразных пара-подий свободно плавают, приступая к половому размножению. Таков знаменитый па-лоло Тихого океана, который периодически в несметном количестве поднимается со дна и роится в поверхностных слоях воды, представляя в это время лакомую пищу для туземцев (Eunice viridis; рис. 222). У других видов задняя половина тела отрывается от передней, регенерирует на месте разрыва новую голову и, обладая более мощными параподиями, уплывает. Бесполая половина остается на прежнем месте и регенерирует задние сегменты. У Autolytus (рис. 223, A), Myrianida и некоторых др. эпитокная половина может еще до отделения от атокной образовать голову, а бесполая—-начать регенерацию задних сегментов: животное имеет вид двух особей разного строения, соединенных зоной регенерации. Иногда, наконец, еще до отделения первого сформировавшегося полового индивида впереди него за счет зоны регенерации образуется вторая, третья и т. д. до 30 половых особей, расположенных в один ряд (рис. 223, А); получается временная цепочка особей (Autolytus и некоторые другие). Затем половые особи отделяются и уплывают (рис. 223, Б, В). Развитие.Дробление яйца полное. Сначала двумя последовательными меридиональными делениями яйцо рассекается на 4 приблизительно одинаковых бластомера: А, В, С и D, причем В отвечает будущей брюшной, a D — будущей спинной стороне зародыша. Затем эти 4 клетки делятся наклонной бороздой на 4 мелкие верхние (анимальные) и 4 нижние крупные (вегетативные) клетки. Получается, как говорят, квартет микромеров и квартет макромеров (рис. 224). Микромеры, смотря по их происхождению от того или иного макромера, обозначают как la, \b, lc
и Id (рис. 224),макромеры—\А — ID. Дальнейшее дробление состоит в последовательном отделении от макромеров по направлению к ани-мальному полюсу вслед за первым еще 2-го, 3-го и 4-го квартетов микромеров, при отделении каждого нового квартета клетки ранее образовавшихся квартетов тоже делятся.
Рис. 223. Размножение полихет. А — бесполое размножение по-лихеты Autolytus (по Грассе); 5 и В — получающиеся посредством концевого почкования половые особи (5 — самка и В — самец) тех же червей (по Свешникову) Важная особенность отделения микромеров заключается в изменении направления веретена деления: анимальный конец его отклоняется то по ходу часовой стрелки, то против нее, если наблюдение вести с ани-мального полюса яйца. Если при образовании 1-го квартета ядерные веретена в бластомерах наклонены в одну сторону, то при отделении 2-го квартета все они оказываются отклоненными в другую, и т. д. Вследствие этого верхние продукты деления ложатся каждый раз не прямо над нижними, а несколько чередуясь с ними, как бы в шахматном порядке. Например, микромер \а лежит не целиком над макромером 1Л, но отчасти налегает и на соседний макромер 1В. Такое дробление называется спиральным, так как если во время деления микромера мысленно продолжить ось ядерного веретена по сферической поверхности яйца, то получится спиральная линия.
Уже на ранних стадиях дальнейшее развитие каждого эластомера точно определено. В общем, первые 3 квартета микромеров идут на образование эктодермы личинки, один из микромеров 4-го квартета — потомок клетки D, обозначаемый 4d, производит целоми-ческую мезодерму, а остальные микромеры 4-го квартета и все макромеры дают энтодерму. Ввиду такого раннего определения дальнейшего развития бластомеров дробление кольчецов называется детерминативным. В результате дробления получается шаровидная бластула, из которой посредством погружения внутрь бла-стоцеля группы крупных эн-тодермальных клеток происходит гаструла. Бласто-пор гаструлы помещается на ее вегетативном полюсе. Затем бластопор вытягивается по одной из сторон гаструлы (будущей брюшной) к ее экватору, а сзади замыкается. Его передняя часть превращается в ротовое отверстие. На нижнем полюсе зародыша образует- ся порошица. Постепенно зародыш превращается в характерную для полихет личинку — трохофору. Трохофора (рис. 225, А)—типичная планктонная личинка, плавающая при помощи ресничек. Тело ее имеет более или менее шаровидную или эллиптическую форму. На переднем (анимальном) полюсе личинки развивается чувствительный теменной султан длинных ресниц, сидящих на группе эктодермальных клеток — теменной пластинке. По экватору личинки впереди рта расположен характерный предротовой венчик ресничек— прототрох. Иногда позади рта развит менее мощный послеро-товой венчик. Кишечник начинается ртом посредине брюшной стороны личинки и заканчивается порошицей на ее заднем полюсе и состоит из трех отделов, причем передняя и задняя кишка образуются впячиванием эктодермы, а средняя кишка формируется из энтодермы. Между кишечником и стенкой тела находится первичная полость тела, пересекаемая тонкими мышечными волокнами. По бокам кишечника лежит пара маленьких протонефридиев. Мезодермальные органы личинки (главным образом мускульные волокна) развиваются из нескольких клеток, лежащих у краев бластопора и называемых мезенхимой. Другой мезодермальный зачаток представлен двумя крупными мезодермальными клетками — первичными мезоб-ластами (иногда их называют телобластами)—потомками бластомера 4d, лежащими по бокам от кишечника. После некоторого периода планктонной жизни трохофоры начинается ее метаморфоз. Заднее (вегетативное) полушарие личинки значитель-
но вырастает в длину и подразделяется сразу на несколько (3, 7, 9—13) сегментов. На сегментах развиваются параподии и щетинки или появляются ресничные пояски. К этому времени обе первичные мезодермальные клетки, усиленно размножаясь, дают два лежащих по бокам от кишечника клеточных тяжа — мезодермальные полоски (рис. 225, Б). Вско-
Рис. 225. Развитие Poltjgordius. A — трохофора, вид сбоку (по Гатчеку); Б — метаморфоз трохофоры (по Вурмбаху): 1 —анус, 2 — задняя кишка, 3 — средняя кишка, 4 — мышцы, 5 — послеротовой и 6 — предротовой венчик ресничек (нрототрох), 7 — теменная пластинка, 8 — теменной султан, 9 — рот, 10—передняя кишка, 11— нижнее полушарие трохофоры, 12—протонефридий, 13 —мезодермальная полоска, 14 — мезобласт, 15 — диссепи- мент, 16 — целом ре под влиянием наружной сегментации мезодермальные полоски расчленяются на парные группы клеток, так что в каждом сегменте оказывается своя пара мезодермальных клеточных зачатков. Последние сначала компактны, затем в них появляется полость — зачаток вторичной полости тела, а ограничивающая ее клеточная стенка есть стенка цело-мического мешка. В каждом сегменте, таким образом, развивается пара целомических мешков. В процессе метаморфоза часть клеток теменной пластинки трохофоры погружается под покровы и образует головной мозг. На брюшной стороне в виде парного валика эктодермы закладываются брюшные нервные стволы. В дальнейшем они вступают в связь с головным мозгом при помощи окологлоточных коннективов. Из эктодермы развиваются и органы чувств — глаза, пальпы. Так, из несегментированной, первичнополостной трохофоры формируется следующая личиночная стадия — метатрохофора (рис. 226), характеризующаяся сегментацией и метамерным целомом. Тело сформированной метатрохофоры состоит из головной лопасти (простомиума), нескольких сегментов и маленькой анальной лопасти (пигидиума) с порошицей на конце. Головная лопасть представляет собой почти неизменившееся переднее предротовое полушарие трохофоры, а анальная лопасть— самый нижний участок трохофоры, лежащий позади уровня первичных мезодермальных клеток. Таким образом, оба отдела метатрохофоры по природе резко отличны от сегментов туловища. Существенная особенность сегментов метатрохофоры заключается в том, что все они возникают одновременно.
Метатрохофора некоторое время плавает или ведет дойный образ жизни, не меняясь существенно, а затем испытывает дальнейший метаморфоз. На переднем крае анальной лопасти образуется зона роста, клетки которой непрерывно размножаются. Область, лежащая впереди нее, состоит из быстро растущих, еще не дифференцированных тканей. В этой зоне формируются новые сегменты и последовательно один за другим отделяются по направлению кпереди (рис. 227). Процесс продолжается до тех пор, пока не образуется столько сегментов, сколько их у взрослого червя. Иногда первичные мезодермальные клетки сохраняются в зоне роста и дают начало мезодермальным полоскам. Чаше, однако, эти клетки целиком расходуются на образование ие-ломической мезодермы метатрохофоры, и мезодермальные полоски образуются за счет размножения эктодермальных клеток зоны роста. От ме-зодермальных полосок последовательно отделяются парные зачатки цело-мических мешков (рис. 227). Каждый вновь образующийся сегмент получает свою пару таких зачатков, из которых развиваются его целомические мешки, разрастающиеся по бокам кишечника кверху и книзу, постепенно вытесняя собой первичную полость тела. Наконец, правый и левый мешки в каждом сегменте встречаются над и под кишкой по спинной и брюшной линиям, а следующие друг за другом пары мешков тоже соприкасаются своими передними и задними стенками
(рис. 228). Наружным кожно-мускульным листком мешки подстилают эктодерму, а внутренним граничат с кишечником. В результате описанного процесса первичная полость тела личинки замещается целомом. За счет соприкасающихся стенок целомических мешков над и под кишкой образуется спинная и брюшная брыжейки, а на месте соприкосно-
Рис. 228. Развитие целома у кольчатых червей. А—В — поперечные разрезы трех последовательных стадий развития сегмента (из Матвеева): / — кишка. 2 — первичная полость тела. 3 — целом, 4 — наружная стенка целомиче- ского мешка, 5 — спинной мезентерий, 6 — внутренняя стенка целомического мешка, 7— брюшной мезентерий, 8 — брюшные нервные стволы вения следующих друг за другом пар мешков — поперечные септы между полостями отдельных сегментов — диссепименты. Продольные кровеносные сосуды — спинной и брюшной — образуются за счет оставшихся участков первичной полости тела внутри брыжеек (между двумя сло-•ями перитонеального эпителия). Производные мезодермы таковы: из кожно-мускульного листка целомических мешков происходит мускулатура тела, из кишечно-мускульно-го — мускулатура кишечника. Кроме того, за счет стенок мешков формируются перитонеальный эпителий и половые воронки (целомодукты). На каждом сегменте развиваются параподии, а внутри него формируются соответствующие участки брюшных нервных стволов, брюшные ганглии, органы выделения и т. п. Важным для понимания организации кольчатых червей, так же как и других типов, берущих от них начало, является двойственность сегментов, входящих в состав тела этих животных. Мы видим, что в процессе метаморфоза сперва образуются сегменты метатрохофоры, затем формируются все остальные (см. рис. 227). Это явление было открыто крупнейшим советским эмбиологом П. П. Ивановым, разработавшим стройную теорию двойственного происхождения метамерии, имеющую большое значение для понимания эволюции всех сегментированных животных. Немногочисленные сегменты метатрохофоры названы П. П. Ивановым ларвальными (т. е. личиночными) в отличие от образующихся позднее постларвальных сегментов. Ларвальные сегменты характеризуются рядом существенных особенностей: образуются одновременно, причем сегментация начинается с наружных органов, отсутствуют половые железы и половые воронки, а иногда отсутствует и метамерность целома. Напротив, при образовании постларвальных сегментов из зоны роста прежде всего сегментируется мезодерма, образуются кровеносная система, половые железы и половые воронки. Таким образом, тело взрослого полимерного червя состоит из следующих различных по происхождению отделов: 1) головной лопасти, или простомиума, представляющего видоизмененное переднее полушарие трохофоры; 2) нескольких ларвальных сегментов; 3) многочисленных постларвальных сегментов и 4) анальной лопасти, или пигидиума, происходящей из самого заднего участка трохофорной личинки. Олигомерные кольчецы (Dinophilus, Myzostornum и др.) в составе туловища имеют только ларвальные сегменты. Поэтому они во многих отношениях напоминают личинку — метатрохофору. Итак, важнейшие черты развития полихет заключаются в спиральном, детерминативном типе дробления, в образовании личинки трохофоры и в закладке мезодермы путем отделения двух мезодермальных полосок от двух мезобластов — первичных мезодермальных клеток; эти клетки называют телобластами, соответственно и данный способ закладки мезодермы именуется телобластическим. Экология.К классу Polychaeta принадлежит около 5300 видов, главным образом морских. Только немногие представители встречаются в пресноводных бассейнах, например в Байкале (Manayunkia). Большинство многощетинковых ведет донный образ жизни, встречаясь главным образом в прибрежной полосе. Многие из них спускаются, однако, глубже 1000 м, а некоторые были найдены даже на глубине 8000 м. Сравнительно немногие виды ведут свободноплавающий образ жизни в планктоне (сем. Alciopidae и др.) и, подобно многим другим планктонным животным, отличаются стекловидной прозрачностью тела. Донные многощетинковые большей частью ползают по дну, среди водорослей, но многие из них ведут роющий образ жизни, проделывая в песке или в иле длинные норы; таков крупный морской червь пескожил — Arenicola (см. рис. 211, Г) и др. Особую биологическую группу составляют сидячие полихеты, выделяющие вокруг себя защитные трубки, из которых высовывается лишь передний конец червя. Размеры представителей класса Polychaeta колеблются в пределах от нескольких миллиметров до 3 м (Eunice gigantea). Палеонтология.Ископаемые остатки полихет немногочисленны. Чаще всего сохраняются трубки сидячих форм, известные еще с докембрия. От бродячих полихет остаются отпечатки щетинок и челюстей, как, например, у Eunicites из литографского сланца Баварии. Практическое значение.Некоторые тропические формы вроде упомянутого выше палоло—Eunice viridis (см. рис. 222) употребляются человеком в пищу. Отдельные виды используются в качестве наживки при ловле рыбы, например пескожил (Arenicola marina', см. рис. 211,Г). Донные формы многощетинковых червей в больших количествах поедаются промысловыми рыбами, камчатским крабом и другими животными. В настоящее время основу пищевого рациона осетровых рыб Каспийского моря составляют Nereis diversicolor— донный многощ-етинковый червь, который отсутствовал в Каспии, но в 1939 и 1940 гг. был переселен туда из Азовского моря. Исследования, связанные с этим успешным переселением ценного кормового объекта, были осуществлены под руководством крупнейшего советского гидробиолога академика Л. А. Зенкевича. Классификация.Класс Polychaeta делится на два подкласса: бродячие (Errantia) и сидячие (Sedentaria).
Поиск по сайту: |