|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
ПОДКЛАСС I. ГИДРОИДНЫЕ (HYDROIDEA)
Строение гидры (Hydra).На примере гидры можно ознакомиться со строением гидроидных полипов. Гидра — одни из наиболее просто устроенных полипов. Это маленький (около 1 см) пресноводный полип, часто встречающийся в озерах и прудах. Тело гидры в виде продолговатого мешочка, прикрепляется к субстрату своим основанием, или подошвой; па свободном конце тела па особом, возвышении — ротовом конусе лежит рот, окруженный венчиком из 6—12 щупалец (рис. 91, А). Вся поверхность тела, вплоть до краев ротового отверстия, покрыта эктодермой, состоящей из нескольких сортов клеток. Большая часть ее образована цилиндрическими или кубическими эпителиальными клетками, основание которых, обращенное к мезоглее, вытягивается по направлению кверху и книзу (по продольной оси животного) в длинный отросток, лежащий параллельно поверхности тела (рис. 91, Г). Цитоплазма отростка дифференцируется в виде тончайших сократительных волоконец; отросток имеет значение мускульного. Цилиндрическая часть клетки входит в состав покровного однослойного эпителия. Эти клетки называются эпителиально-мускульными. Совокупность отростков всех таких
клеток образует в основании эпителия слой мускульных образований, совпадающих с продольной осью тела. При их одновременном сокращении тело полипа сильно укорачивается. Между основаниями более крупных эпителиально-мускульных клеток располагаются мелкие промежуточные (интерстициальные) клетки. За их счет формируются половые и стрекательные клетки. Непосредственно под эпителием рассеяны нервные клетки звездчатой формы, которые своими отростками сообщаются между собой и образуют субэпителиальное нервное сплетение. Таким образом, нервная система гидры стоит на самой низкой ступени развития, имеет рассеянный, диффузный характер (рис. 92). Впрочем, даже у гидры отмечаются два сгущения нервного сплетения — вокруг рта и на подошве. Характерная черта кишсчнополостных — присутствие в покровах стрекательных клеток (рис. 93, А, Б). Они развиваются из промежуточных клеток и содержат особую овальную стрекательную капсулу с плотными стенками. Капсула наполнена жидкостью, а на одном конце капсулы стенка ее впячена внутрь в виде очень тонкого, но полого отростка, который закручивается в капсуле в спирально завитую стрекательную нить. Стрекательные клетки служат гидре орудием нападения и защиты.
Рис. 91. Гидра Hydra oligactis. A—продольный разрез; Б — поперечный разрез; В — участок среза при большом увеличении; Г — эктодермальная эпителиально-мускульная клетка (А — из Бриана, Б — по Полянскому, В — по Кестнеру, Г — по Роскину): / — эктодерма, 2 — энтодерма, 3 — базальная мембрана, 4 — гастральная полость, 5 —эктодер-мальные эпителиально-мускульные клетки, 6 — интерстициальные клетки, 7 — стрекательные клетки, 8 — нервные клетки, 9 — энтодермальные " эпителиально-мускульные клетки, 10 — железистые клетки, // — ротовое отверстие, 12 — ротовой конус, 13 — почка, 14 — подошва, 15 — яйцеклетка, /6 — мужские гонады . 117
На наружной поверхности клетки имеется тонкий чувствительный волосок— книдоциль. Изучение стрекательных клеток с помощью электронного микроскопа показало значительную сложность строения книдоциля (рис. 93, В). Он состоит из длинного жгутика, окруженного 18—22 тонкими пальцевидными выростами цитоплазмы — микроворсинками. По строению жгутик книдоциль очень сходен со жгутиками и ресничками простейших, но в отличие от них неподвижен. При прикосновении добычи или врага к жгутику последний отклоняется и задевает одну или несколько микроворсинок, что приводит к возбуждению стрекательной клетки. При этом стрекательная капсула выбрасывает выворачивающуюся из нее наружу упругую нить, которая распрямляется, как стрела. Нить наподобие гарпуна усажена обращенными назад шипиками, а в основании несет более крупные шипы. Уколы нити ядовиты и могут парализовать мелких животных. После выбрасывания нити стрекательная клетка погибает. У гидры имеется несколько категорий капсул, функции которых различны. Рассмотренные крупные капсулы, служащие для пробивания покровов и поражения добычи, называются пенетрантами (рис. 93). Значительно более мелкие — вольвенты имеют короткие спирально закрученные нити, которые обвиваются вокруг различных выступов (щетинок, волосков и т. п.) на теле добычи и таким путем удерживают ее. Наконец, вытянутые стрекательные капсулы — глютинанты — приклеиваются к телу добычи длинными липкими нитями.
Энтодерма выстилает всю гастраль-ную (пищеварительную) полость вплоть до краев рта. В состав энтодермы входит также несколько кате-
Рис. 93. Стрекательные клетки. А — в покоящемся состоянии; Б — с выброшенной стрекательной нитью (по Кюну); В — строение книдоциля (по Слаутербаку): 7 — ядро, 2 — стрекательная капсула, 3 — книдоциль, 4 — стрекательная нить с шипиками, 5 — шипы, 6 — жгутик, 7 — микроворсинки горий клеток (см. рис. 91, В). Основу ее составляют эпителиально-мускульные пищеварительные клетки, кроме них имеются особые железистые клетки. Мускульные отростки энтодермальных эпителиально-мускульных клеток расположены поперечно по отношению к продольной оси тела. При сокращении их тело гидры суживается, становится тоньше, т. е. они антагонисты эктодермальных эпителиально-мускульных клеток. Эпителиальная часть энтодермальных клеток, направленная в сторону пищеварительной полости, несет 1—3 жгута и способна образовывать псевдоподии, которыми захватываются мелкие пищевые частицы. Таким образом, у кишечнополостных, как и у губок, имеет место внутриклеточное переваривание пищи — признак примитивной организации. Однако наряду с этим железистые клетки энтодермы выделяют пищеварительные соки непосредственно в гастральную полость, где также происходят процессы пищеварения, т. е. у гидры сочетаются внутриклеточное и полостное пищеварение. Гидра питается различными мелкими животными, преимущественно рачками (дафнии, циклопы). Мезоглея у гидры и других гидроидных полипов в виде тонкой бесструктурной пластинки — базальной мембраны, залегающей между эктодермой и энтодермой. Размножение и развитие.Гидры размножаются бесполым и половым путями. Бесполое размножение состоит в почковании. Приблизительно на уровне середины тела гидры имеется так называемый пояс почкования. Здесь время от времени образуется бугор (почка), вырастающий и образующий на вершине новый рот и зачатки щупалец. Почка у основания перешнуровывается, падает на дно и переходит к самостоятельной жизни. Иногда почкование идет так энергично, что еще до отрывания первой почки на гидре успевают образоваться две-три другие (рис. 94). Половым способом гидра начинает размножаться с приближением холодов. Гидры раздельнополы, но встречаются и гермафродитные виды, причем половые клетки их происходят из эктодермы, что характерно для всех Hydrozoa. Некоторые интерстициальные клетки эктодермы или непосредственно превращаются в яйца, или же многократным делением
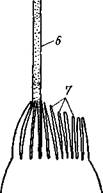
Рис. 94. Hydra oligactis при небольшом увеличении. А — с мужскими гонадами; Б — с женскими гэнадами; В — почкующаяся гидра (по Полянскому) дают скопления живчиков (сперматозоидов). В этих местах на теле гидры эктодерма вздувается в виде бугорков. Яйца располагаются ближе к основанию гидры, а бугорки со сперматозоидами (мужские гонады) — к ротовому полюсу. Яйцо оплодотворяется в теле матери еще осенью и окружается плотной оболочкой, потом гидра погибает, а яйца остаются в покоящемся состоянии до весны, когда из них развиваются новые гидры. Морские гидроидные полипы.Лишь очень немногие гидроидные полипы наподобие гидры ведут одиночный образ жизни. Большинство образует колонии, состоящие из множества особей (рис. 95). Формирование колоний становится легко понятным из сравнения с гидрой. Представим себе, что почки, образовавшиеся на теле гидры, не отрываются от нее, а остаются с ней в постоянной связи и сами начинают почковаться, не отделяя от себя образующихся дочерних полипов. Получается комплекс особей, сидящих как бы на общем стволе и его побочных ветвях. Колония чаще всего имеет вид деревца или кустика. Основание общего ствола колонии обыкновенно дает стелющиеся по субстрату отростки, похожие на корни растений и служащие для прикрепления колонии. Ствол ветвится, на ветвях сидят отдельные особи колонии — гидранты;
Рис. 95. Гидроид Obelia. A — колония (слегка увеличено); Б — отдельная веточка колонии (несколько схематизировано, часть особей колонии изображена в разрезе) (по Абрикосову): 1 — гидрант в расправленном состоянии, 2 — сократившийся гид
каждая особь соответствует как бы одной почке гидры и напоминает гидру. Гистология гидранта в общем отвечает таковой гидры, но гаст-ральная полость гидрантов продолжается в полость, или капал, проходящий через весь ствол и ветви колонии. Таким образом, гастральные полости всех гидрантов сообщаются между собой, так что пища, захваченная отдельными гидрантами, может затем в переработанном виде распределяться по всей колонии. Эктодермальный эпителий ствола выделяет на поверхности особую
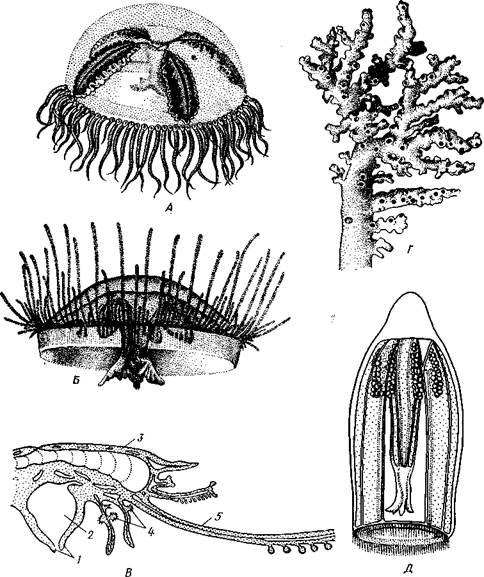
Рис. 96. Сравнение строения гидроидного полипа (Л) и гидроидной медузы (Б), перевернутой ротовым отверстием кверху (по Холодков- скому): I — рот, 2 — щупальца, 3 — гас-тральная полость, 4—мезоглея, 5 — радиальный канал, 6 — парус Рис. 97. Гидроидная медуза Sarsia (рис. А. Наумова): J — рот, 2 — ротовой стебелек с расположенной на нем гонадой (3), 4 — радиальные каналы, 5 — кольцевой канал, 5 — щупальца, 7 — глазки, 8 — парус органическую оболочку — теку, которая придает ему большую устойчивость. Тека доходит до основания гидрантов (подотряд Athecata), а иногда продолжается и на них самих (подотряд Thecaphora) в виде защитного колпачка, или гидротеки. Размножение морских гидроидных полипов, образование и строение медуз.Сами полипы способны лишь к бесполому размножению почкованием, в гидрантах половые железы не образуются. Они формируются лишь в специальных, возникающих на колонии тоже путем почкования половых особях, медузах, переходящих к свободноплавающему образу жизни. Сначала на известных местах стебля колонии появляется вырост, напоминающий зачаток гидранта. Этот вырост сильно вытягивается и превращается в полый столбик, видоизмененный полип — бластостиль, по бокам которого выпочковываются зачатки медув. Каждый зачаток постепенно превращается в молодую медузу, которая отрывается от бла-стостиля и уплывает. Медузка растет, развивает в себе половые клетки и приступает к половому размножению. Иногда медузы выпочковываются поодиночке, без бластостиля. Медузы, за исключением некоторых отклонений в организации пищеварительной системы, построены по той же схеме, что и полипы, но часто сильно сплющены в плоскости, перпендикулярной к главной оси тела (рис. 96). Медуза имеет вид колокола или зонтина; наружная выпуклая сторона называется эксумбреллой, внутренняя вогнутая — субумбреллой (рис. 97). Посредине последней выдается более или менее длинный ротовой стебелек со ртом на свободном конце. Рот ведет в пищеварительную, или гастральную, полость, состоящую из центрального желудка и расходящихся от него к краям зонтика радиальных каналов в числе, равном четырем или кратном четырем, и соединенных в толще мезоглеи сплошной энтодермальной пластинкой. На краю зонтика все радиальные каналы сообщаются между собой посредством кольцевого канала. Желудок и каналы в совокупности образуют гастроваскулярную (т. е. кишечносо-судистую) систему. По свободному краю зонтика прикреплена тонкая кольцевидная мускулистая перепонка, суживающая вход в полость колокола. Она называется парусом и является характерной особенностью гидроидных медуз, отличающей их от медуз, принадлежащих к Scypho-zoa. Парус играет важную роль при движении медуз. На краю зонтика расположены, кроме того, щупальца. Они, подобно радиальным каналам, имеются в определенном числе, чаще всего кратном четырем. Вследствие правильного расположения радиальных каналов и щупалец лучистая симметрия медуз ярко выражена. Тело медуз характеризуется сильным развитием мезоглеи, которая очень утолщается и содержит большое количество воды, приобретая студенистый желеобразный вид. Благодаря этому все тело медуз почти стекловидно и прозрачно. Прозрачность, свойственная очень многим планктонным животным, рассматривается как особый род покровительственной окраски, укрывающей животное от врагов.
Нервная система медуз устроена значительно сложнее, чем у полипов. У медуз кроме общего подкожного нервного сплетения по краю зонтика наблюдаются скопления ганглиозных клеток, которые вместе с отростками образуют сплошное нервное кольцо. От него иннервируются мышечные волокна паруса, а также особые органы чувств, расположенные по краю зонтика. У одних гидроидных медуз эти органы имеют вид глазков, у других — так называемых статоцистов, или органов равновесия (рис. 97, 98). Глаза медуз в наиболее примитивном виде устроены по типу простых глазных пятен. У основания некоторых щупалец имеется небольшой участок эктодермального эпителия, состоящий из клеток двух родов. Одни из них высокие — чувствительные, или ретипальные, клетки; другие содержат многочисленные бурые или черные зерна пигмента и чередуются с чувствительными клетками, согюкупность которых отвечает сетчатке глаза высших животных. Присутствие пигмента вообще характерно для органов зрения во всем животном царстве. Несколько более сложно устроены глазные ямки, где пигментированный участок эпителия лежит на дне небольшого ппячивания покрова. Подобный уход глаза с поверхности тела вглубь предохраняет его от различных чисто механических раздражений, например трения о воду, прикосновения посторонних предметов и т. п. Кроме того, впячивание глаза ведет к увеличению поверхности светочувствительного слоя и количества ретинальных клеток. Наконец, у некоторых медуз полость глазной ямки заполняется прозрачным выделением эктодермы, получающим вид лучепрсломляющей чечевицы (линзы). Таким путем возникает хрусталик, концентрирующий световые лучи на сетчатке глаза. Органы равновесия могут быть устроены различно: в виде чувствительных щупалец, но чаще всего в виде глубоких эпителиальных ямок, которые могут отшнуровываться от поверхности тела и превращаться в замкнутые пузырьки, или статоцисты (рис. 98). Пузырек выстлан чувствительным эктодермальным эпителием и заполнен жидкостью. Одна из клеток пузырька впячивается внутрь него в виде вздутой на конце булавы, внутри которой выделяется одна или несколько конкреций углекислой извести. Это статолиты, или слуховые камешки, и они столь же характерны для органов чувства равновесия, как пигмент для органов зрения. Чувствительные клетки пузырька снабжены каждая длинным чувствительным волоском, направленным к находящейся в центре его булаве. Строение волоска сходно со строением книдоциля стрекательных клеток. По функции статоцисты медуз более или менее отвечают функции полукружных каналов уха человека. Новейшие электронно-микроскопические исследования показали, что волоски чувствительных клеток в статоцистах медуз построены по одному типу с чувствительными волосками рецепторных органов более высоко организованных животных вплоть до позвоночных. В последнее время статоцисты медуз считают не только органами равновесия, но и приспособлениями, стимулирующими сократительные движения краев зонтика: если вырезать у медузы все статоцисты, то она перестанет двигаться. Медузы раздельнополы. Половые железы (гонады) располагаются у них на нижней стороне зонтика, под радиальными каналами или на ротовом стебельке, и представляют собой кучки половых клеток, лежащих между эктодермой и мезоглеей. Медузы плавают в толще воды, частью переносимые морскими течениями, частью двигаясь активно при помощи действия мышечных волокон, имеющихся по краю зонтика и в парусе. Одновременным сокращением зонтика и паруса и их последующим расслаблением вода, которая находится в вогнутости зонтика, то выталкивается из него, то пассивно вновь его заполняет. При выталкивании воды животное получает обратный толчок и двигается выпуклой стороной зонтика вперед. Вследствие чередования сокращений и расслаблений зонтика и паруса движение медуз состоит из ряда прерывистых толчков. Медузы — хищницы. Своими щупальцами они захватывают и убивают различных мелких животных, проглатывают их и переваривают в га-стральной полости. Половое размножение и развитие.Половые клетки после созревания выходят наружу через небольшие разрывы наружной стенки тела. Опло-
Рис. 99. Развитие медузы Aequorea (по Клаусу). Гаструляция происходит посредством иммиграции клеток бластулы (А) на вегетативном полюсе последней (5, В), в результате получается двухслойная личинка планула (Г) дотворенне и все дальнейшее развитие яиц протекает вне материнского организма. Яйцо испытывает полное равномерное дробление, в результате которого формируется продолговатая бластула (рис. 99). Переход от бластулы к двухслойному состоянию — гаструляция — у громадного
большинства кишечнополостных совершается не впячиванием одной половины бластулы в другую, а путем иммиграции. На заднем полюсе бластулы часть клеток ее стенки начинает поодиночке уходить внутрь личинки, т. е. в блас-тоцель. В конце концов полость заполняется сплошной массой однородных клеток, которые представляют зачаток энтодермы. Развившаяся таким образом личинка — паренхимула — напоминает паренхимулу губок (с. ПО). В дальнейшем часть эн-тодермальных клеток личинки дегенерирует и на их месте возникает небольшая полость — зачаток будущей гастральной полости. На этой стадии, характерной для кишечнополостных, личинка называется планулой (рис. 99).
Планула овальной формы, сплошь покрыта ресничками, при помощи которых она некоторое время свободно плавает, а потом опускается на дно и прикрепляет- ся к нему слегка расширенным передним концом. В энтодерме осевшей планулы формируется гастральная полость; на конце, противоположном месту прикрепления, прорывается отверстие, сообщающее эту полость с внешней средой, — ротовое отверстие, по краям последнего вырастает венчик щупалец и планула превращается в маленького полипа. Полип растет, почкуется и дает колонию гидроидных полипов.
Чередование поколений.Итак, жизнь гидроидных полипов состоит из правильного чередования двух поколений, различных по строению и способу размножения. Одно поколение, полипоидное, ведет сидячий образ жизни и размножается только бесполым способом, производя посредством почкования полипов и медуз. Медузы (второе поколение) отрываются от колоний полипов и переходят к свободному подвижному образу жизни. Они размножаются половым путем и снова дают начало поколению полипов. Такое чередование поколений, размножающихся различным способом, а именно половым и бесполым, называется метагенезом (рис. 100).
Большинство Hydroidea имеет типичное чередование поколений. У некоторых представителей, однако, наблюдается уклонение от намеченной схемы, сказываю- щесся в частичном подавлении одного из поколений, а именно медузоидного. Образующиеся на колонии медузы перестают отрываться от нее и, оставаясь на месте, т. е. на бласто-стиле, развивают в себе половые клетки. Такие медузы, или медузои-ды, отличаются недоразвитием рта, органов чувств и некоторых других органов. Подавление медузоидного поколения может идти еще дальше, причем медузы постепенно утрачивают характерную форму и превращаются в простые, набитые половыми клетками мешки (гонофоры), сидящие на колонии полипов (рис. 101). Будучи сначала свободнопод-вижным самостоятельным поколением, медузы, таким образом, постепенно становятся как бы половыми органами колонии полипов: интересный пример низведения особи до степени простого органа. Но иногда изменение в смене поколений идет, по-видимому, в обратном направлении, причем медузоидное поколение получает полное преобладание над полипоидным. По крайней мере, подобно тому как мы встречаем среди гидрозоев формы чисто полипоидные (гидра), не дающие никогда медуз, точно так же есть группа медуз (отр. Trachylida), которые в течение всего жизненного цикла остаются медузами. Они похожи на описанных раньше гидроидных медуз; планула этих медуз не
оседает на дно, но превращается прямо в медузу. Таким образом, здесь чередование поколений исчезает и остается только одно поколение — медузоидное. Классификация. Отряд 1. Leptolida— формы с различно выраженным чередованием поколений и в полипоидной стадии обычно образующие кустистые или древовидные колонии. В отряде четыре подотряда: Alhecata, Thecaphora, Limnomedusa и Hydrocorallia. Это самая многочисленная группа гидроидных. Колонии подотрядов Athecata и Thecaphora (см. рис. 95) образуют густые заросли на морском дне, водорослях, ракуш-
Рис. 102. Различные Hydrozoa. А — ядовитая медуза «Крестовичок» (Gonionemus); £ — пресноводная медуза Craspedacusta; В — схема строения Porpita — представителя отряда Chondrophora; Г—известковая колония Hydrocorallia; Д — трахимедуза Aglantha (по Наумову, В — из Байера): / — ротовое отверстие, 2 — кишечная полость. 3 — хитиноидная скелетная пластинка, 4 — гонофоры, 5 — щупальце ках, сваях и т. п. Отдельные гидранты очень мелкие, чаще не более 1 мм, только некоторые Athccata, например Tubularia (рис. 101), достигают нескольких сантиметров, а обитающий у берегов Японии одиночный полип Branchiocerianthus—даже до 1 м. Медузы обычно тоже небольшой величины, от 5 мм до 2—3 см в диаметре зонтика, редко бывают крупнее. Большинство представителей отр. Leptolida обитает в море, п пресных водах встречается лишь несколько видов подотряда Limnomedusae, характеризующегося преобладающим развитием медузоидного поколения. Так, в пресноводных водоемах субтропиков и тропиков всех материков, а также в аквариумах с тропическими растениями и рыбками периодически появляются медузы Craspedacusta (рис. 102, Б). К, лимпомеду-зам относится также очень ядовитая морская медуза «крестовичок» (Gonionemus), обитающая у берегов Японского моря и Курильских островов (рис. 102, А). Полипы этих видов едва заметны простым глазом и не образуют колоний. Представители Hydrocorallia (рис. 102, Г) отличаются массивным известковым скелетом, напоминая этим некоторых коралловых полипов (с. 140). Встречаются в умеренных и тропических морях. Отряд 2. Chondrophora— крупные одиночные полипоидные формы, обитающие па поверхности воды тропических морей. Тело уплощенное, рот, щупальца и гонофоры обращены вниз (рис. 102, В). На аборалыгой стороне имеется хитиноидная пластинка, соответствующая теке других гидроидных. У Vclella (табл. V, 2) она выступает вверх о виде треугольного паруса. Отряд 3. Trachylida— чисто мсдузоидные формы, не имеющие смены поколений. Относящаяся сюда Aglantha (рис. 102, Д) встречается по всему Мировому океану, другой представитель — Сипта паразитирует на медузах отряда Leptolida. Вообще среди кишечнополостных насчитывается очень немного паразитических форм. Особый интерес среди них представляет вид Polypodium hydriforme, систематическое положение которого до сих пор не установлено. Polypodium паразитирует на яйцах осетровых рыб. В икринке хозяина Polypodium имеет вид длинного извитого тяжа— столона, образующего на себе ряд почек со щупальцами. Интересно, что внутри икринки почки Polypodium находятся в вывернутом наизнанку состоянии: эктодерма его обращена внутрь, а энтодерма — наружу к питательному желтку яйца. Позднее почки выворачиваются эктодермой наружу и, когда икра отложена в воду, столон расч издается; образуется 60—90 полипов, которые переходят к свободной жизни, прикрепляясь ко дну. В течение лета полипы размножаются продольным делением. К осени на них образуются половые железы. Дальнейшая судьба полипа и способ заражения хозяина неизвестны. Отряд 4. Hydrida— одиночные формы, чисто полипоидные, без чередования поколений. Представитель — пресноводная гидра Hydra и немногие другие. В ископаемом состоянии гидроидные известны, начиная с силура. Главным образом это массивные известковые Hydrocorallia. Имеются также отдельные сведения о нахождении ThecapKiora, уже начиная с кембрия.
Поиск по сайту: |