|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
Некоторые нейроанатомические проблемы восприятия
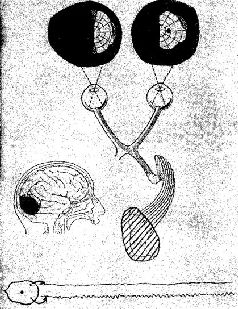
К числу главных факторов, препятствующих пониманию неврологии процесса восприятия, относится одно из тех противоречий, один из тех парадоксов, с которыми мы неоднократно встретимся в этих главах, пытаясь связать функции нервной системы с психологическими процессами. В данном случае парадокс касается неироанатомическои организации системы входа и тех нарушений в поведении, которые возникают в результате разрушения этой организации (см. Рис. VII—1).
В сущности, анатомическая организация систем входа является единственной организацией, в которой сохраняется топологическое соответствие между рецепторной поверхностью и корой головного мозга. Это означает, что расположение точек на сетчатке будет проецироваться на кору в виде сходного расположения точек. Таким образом могут быть построены карты ретино-кортикального соответствия. Такие карты строятся на основе данных, полученных либо методом дегенерации нервных волокон после небольших разрушений в рецепторных системах (Browner, 1934; Browner and Zeeman, 1926), либо в результате точечной электрической или световой стимуляции сетчатки при одновременной регистрации электрических ответов от участков ‹коры, в которых они могут быть вызваны (Talbot and Marshall, 1941). Подобные карты были пелучены для кожной чувствительности. По этому же принципу устроен и кохлеарно-кортикальный слуховой механизм (Rose and Woolsey, 1949). Такая рецепторно-кортикальная организация не сопровождается прямым однозначным соответствием посредством единой нервной нити. Скорее всего, столь же сложная организация, которую мы уже анализировали на уровне сетчатки, повторяется как на каждом уровне механизма рецепторной системы, так и в коре. Фактически наблюдается общее сокращение числа клеток в ганглиозном слое сетчатки по сравнению с рецепторным (у человека это уменьшение оценивается примерно как 100 или 50 к одному). При переходе от слоя ганглиозных клеток к латеральному коленчатому телу таламуса число зрительных элементов, по существу, остается постоянным. При переходе от таламуса к коре возникает обратная картина: одиночная.клетка латерального коленчатого тела может контактировать с 5000 кортикальных нейронов каждый из которых находится в контакте еще примерно с 4000 других нейронов через их дендритные связи. Благодаря такому механизму тормозных взаимодействий при стимуляции у обезьяны двух точек сетчатки в фовеа, несмотря на некоторое перекрытие зон возбуждения, возникает их раздельное восприятие, причем таким образом, что двум угловым минутам на сетчатке соответствует расстояние в 1 мм на поверхности коры (Talbot and Marchall, 1941, p. 134). Казалось бы, что такое строение нервной системы обеспечивает проекцию некоторого «образа» от рецепторпой поверхности на поверхность коры во многом так же, как проецируется изображение в кинокамере. Парадокс проявляется, когда в результате болезни или хирургической операции повреждаются системы входа. Правда, как и ожидалось, в некоторых случаях может быть обнаружено выпадение части поля зрения (скотома) в месте, которое может быть предсказано в соответствии с анатомической организацией мозга (см. Рис. VII—2).
Однако при сохранении хотя бы малой части входного механизма это выпадение часто не замечается даже в том случае, если глаза удерживаются в неподвижности. При этом распознавание структуры сохраняется и во многих отношениях не отличается от нормального. Люди с громадными скотомами либо совсем не знают о них, либо могут быстро научиться легко их игнорировать. Животное, у которого было удалено или разрушено от 80 до 98 процентов нервных связей входного механизма, способно решать задачи, требующие различения структур, отличающихся только деталями. Лешли (1929) удалял у крыс 80—90 процентов стриарной коры, не нарушая их способность различать зрительные структуры. Р. Галамбос перерезал у кошек 98 процентов волокон зрительного тракта, и животные еще могли хорошо выполнять задания, требовавшие различения весьма сходных фигур (Galambos, Norton and Frommer, 1967). Недавно Чау (1970), также экспериментируя с кошками, разъединял более 3/4 волокон зрительного тракта и удалял более 3/4 зрительной коры; в таких условиях едва ли оставалась неповрежденной любая из проекционных систем, характеризующихся точечным («точка в точку») представительством. Хотя зрительное различение структур после такого значительного разрушения вначале оказалось нарушенным, животное снова научилось выполнять задание, на что ему потребовалось примерно такое же число проб, как и до операции. В моих экспериментах, как связанных с клинической нейрохирургией, так и проведенных в лаборатории (например, Wilson and Mishkin, 1959), удаление мозговой ткани было локальным и ограничивалось корой, так что оно не вызывало массивного разрушения белого вещества, благодаря чему у пациента или подопытного животного сохранялась в значительной мере способность к восприятию. После временно возникающей скотомы, длящейся несколько недель, можно было обнаружить лишь очень незначительные нарушения зрительной функции. Как уже отмечалось, исследователи пытались использовать и многие другие методы для разрушения предполагаемой ими организации входных систем мозга, но безуспешно: Р. Сперри и его группа (1955) хирургически производили перекрестное рассечение области, воспринимающей сенсорные сигналы, и даже помещали полоски слюды в места рассечения, чтобы электрически изолировать маленькие квадратики ткани друг от друга. Лешли, Чау и Семмез (1951) пытались накоротко замкнуть электрическую активность мозга, накладывая полоски золотой фольги на воспринимающие поверхности. Я создавал в воспринимающей области коры множество эпилептогенных очагов посредством инъекции микроскопического количества алюминиевой пасты (Kraft, Obrist and Pribram, 1960; Stamm and Pribram, 1961; Stamm and Warren, 1961). Хотя такое множество очагов заметно задерживало научение различению зрительных структур, оно не препятствовало его проявлению, если научение уже закончилось или происходило после того, как были произведены многочисленные разрушения. Эти результаты показывают, что эффект входного сенсорного воздействия на мозговую ткань, получаемая на входе информация распределяются по всей входной системе. Запись электрической активности мозга также свидетельствует о том, что в мозгу происходит распределение информации. Э. Р. Джон (John, Herrington and Sutton, 1967), например, при изучении зрительной системы использовал технику «меченых стимулов», предъявляя кошкам стимулы, которые различались не только своей геометрической структурой, но и частотой образующих эти стимулы мельканий. Эта различная частота мельканий отражалась на электрической активности мозга в ответ на предъявление стимула (или, если это мелькание было очень быстрым, то возникала субгармоника этой частоты). Таким образом, кодируемое различие частоты может оставить «след» в мозгу. Эта техника дала ряд интересных результатов, но для нас здесь важнее всего то, что тщательный анализ меченых волновых форм (вычисление позволяет обнаружить различие в волновых формах, возникающих в различных участках мозга) показывает, что одинаковые меченые волновые формы возникают во многих структурах мозга одновременно.
Однако другая группа экспериментов, проведенных в моей лаборатории (Pribram, Spinelli and Kamback, 1967, Pис. VII—3, VII—4), показывает, что если научение уже имело место, то информация в мозгу не распределяется по всей системе. Были использованы очень маленькие электроды. Обезьяны были обучены по-разному реагировать на различные геометрические стимулы. В отличие от экспериментов Джона стимулы освещались очень короткой вспышкой света В зрительной коре было обнаружено несколько различных типов волновых форм электрической активности. Один тип был получен, когда форма волны определялась относительно момента предъявления стимула. Этот тип вызванного ответа обнаружил четкую зависимость от характеристики стимула. Два других типа были получены, когда форма волны определялась по отношению к началу ответа животного. Один тип волновой формы зависел от того, получила обезьяна подкрепляющий пищевой шарик за правильный ответ или не получила, когда ее ответ был ошибочным Другой тип волновой формы возникал непосредственно перед двигательной реакцией животного. Эта форма волны коррелировала с определенным типом двигательного ответа (нажатием на правую или левую панель); она следовала за стимулом, но не зависела от него и от полученного подкрепления. Важным для нас является тот факт, что все эти типы характерных волновых форм появляются локально в зрительной коре. Один тип волновой формы записан от одних электродов, другой от других. Распределение этих волновых форм происходило не в соответствии с какой-либо структурой возбуждения. Однако со дня на день, от недели к неделе наблюдалось полное постоянство в записях, получаемых от каждого отдельного электрода. Какой бы процесс кодирования ни происходил, он стабилизировался по мере того, как проводилась запись.
Результаты этих экспериментов не согласуются с представлением о том, что образ, подобно фотографии, проецируется на поверхность коры. Они показывают, что каждая сенсорная система функционирует, имея большой резерв. Поскольку системы с разрушением и без него обнаруживают лишь незначительные различия в поведении, этот резерв, должно быть, распределен по всей системе — сохраняемая в ней информация, необходимая для осуществления различения, дублируется одновременно многими ее участками. Таким образом, вполне вероятно, что задержка в научении, возникающая при наличии эпилептогенных очагов, вызванных наложением на поверхность мозга алюминиевой пасты, обусловлена помехами, которые вносятся этими очагами в процесс дублирования хранения информации (см. Рис. VII—5). Вопросы, поднятые этими наблюдениями, следует сопоставить с другим рядом проблем: каким образом предметы кажутся нам достаточно стабильными, так что мы можем узнавать их независимо от угла зрения и расстояния до предмета? Каким образом мы узнаем предмет, несмотря на то, что светом, идущим от предмета, возбуждается только часть сетчатки, а следовательно, и мозга? Константность восприятия предметов и величины существует у детей уже в возрасте нескольких недель. Таким образом, простое объяснение феномена константности восприятия в терминах научения ставится под сомнение. Какого же рода механизм мог бы обеспечить одновременно существующую гибкость восприятия и константность узнавания?
Как факты восприятия структур при наличии скотомы, так и факты константности восприятия говорят о том, что существует, должно быть, какой-то эффективный нейрологический механизм пространственного распределения информации на входе мозга. Для объяснения фактов восприятия недостаточно изоморфизма, простого («точка в точку») соответствия между стимулом и картиной возбуждения в коре, обусловленной анатомией системы. Когда в результате удаления коры происходит выпадение 80 процентов зрительного поля, узнавание осуществляется благодаря оставшейся части зрительного поля; когда в зрительной коре производится множество разрушений, то сохранившиеся части функционируют столь успешно, что при различении испытываются лишь незначительные трудности; рассматриваем ли мы предмет той или другой частью сетчатки, под тем или иным углом зрения, мы всегда можем узнать его. Восприятие отличается от обычного фотографического изображения — оторвите 98 или даже 80 процентов у фотографии и попытайтесь идентифицировать ее: в большинстве случаев это невозможно.
Поиск по сайту: |