|
|
|
Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
КИНЕТИКА ВОСПАЛИТЕЛЬНОГО ПРОЦЕССА ⇐ ПредыдущаяСтр 2 из 2
Сопряжение воспаления с иммунитетом для репарации обес печивается участием всех систем защиты организма в уникал ной реакции терминальных сосудов и соединительной ткан„, которая составляет сущность воспаления. Как известно, защиту организма определяют неспецифические факторы и иммунологическая реактивность, или иммунный ответ. Неспецифические факторы защиты и иммунологическая реактивность [по Петрову Р.В., 1982]
В становлении иммунитета при воспалении велика роль ка фагоцитоза, так и системы комплемента. Место фагоцитоза, осуществляемого полиморфно-ядерными лейкоцитами (ПЯЛ) и мо-ноцитарными фагоцитами (макрофаги), в системе иммунитета определяется тем, что, несмотря на неспецифичность самого акта фагоцитоза, фагоциты, особенно макрофаги, принимают участие в очищении антигенов, переработке их в иммуногенную форму, которую воспринимает Т-хелпер. Место макрофагов системе иммунитета определяется и участием в кооперации Т- и В-лимфоцитов, необходимой для становления иммунного ответа. Поэтому фагоцитоз дополняет формы реакций иммунологической реактивности. Система комплемента участвует в специфических реакциях, присоединяя свои компоненты к молекулам антител, что обеспечивает лизис антигенных субстанций, против которых выработаны антитела. Из этого следует, что комплемент как один из неспецифических факторов защиты принимает участие в иммунном ответе, поэтому он, как и фагоцитоз, дополняет формы иммунологической реактивности. Как видно, включение иммунного ответа при воспалении обеспечивается двумя клеточными системами неспецифической защиты: системой моноцитарных фагоцитов, а также плазменной системой — системой комплемента. Кинетика воспалительной реакции для достижения конечной цели — элиминации повреждающего агента и репарации ткани — характеризуется сменой взаимоотношений клеточных систем защиты между собой и с системой соединительной ткани, что определяется медиаторной регуляцией. Из этого, однако, не следует, что в воспалительной реакции участвуют лишь ПЯЛ, макрофаги, лимфоциты и фибробласты. Клетки — носители вазоак-тивных аминов (лаброциты, базофилы, тромбоциты), как и сдерживающие их функциональную активность эозинофилы, имеют огромное значение для развития собственно сосудистой реакции воспаления. Но они непричастны к основному назначению воспалительной реакции — элиминации повреждающего начала и репарации повреждения. Как цепная, в значительной мере саморегулирующаяся, воспалительная реакция укладывается в универсальную схему: повреждение -+ медиация ► рецепция ► кле точная кооперация > клеточные трансформации ► репарация (схема 16). Воспалительная реакция определяет и последовательно развивающиеся фазы: 1) повреждения, или альтерации, 2) экссудации, 3) пролиферации и дифференцировки клеток. Повреждение (альтерация) — обязательный компонент воспаления. Это изначально то, на что возникает со-судисто-мезенхимальная реакция, составляющая сущность воспаления. Можно ли считать альтерацию фазой воспаления? Вопрос этот не решается однозначно. Одни современные патологи IRobbins S. et al., 1981] не выделяют альтерацию как таковую, подменяя ее нарушениями микроциркуляции и реологических свойств крови. А.М.Чернух в монографии "Воспаление" (1979) первой стадией воспаления называет сосудистую, выделяя в ней две фазы. Д.С.Саркисов и В.Н.Галанкин (1988) рассматривают альтерацию как неспецифический компонент воспаления, причем не всегда обязательный (В.Н.Галанкин) для развития после-
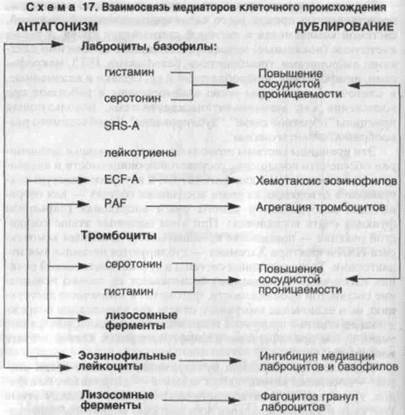
дующей экссудации и пролиферации. Иными словами, допускается возможность развития воспаления без повреждения, причем альтерация в такой ситуации подменяется функциональной недостаточностью полиморфно-ядерных лейкоцитов. Эта позиция, даже условно допускаемая, исключает понимание воспаления как сосудисто-мезенхимальной реакции на повреждение. Многие патологи [Струков А.И., 1972; Серов В.В., Пауков B.C., 1995; Cottier H„ 1980] отстаивают необходимость выделения альтеративной фазы воспаления, характеризующей инициальные процессы (дистрофия, некроз) и выделение медиаторов. Вероятно, у патологоанатома есть все основания для сохранения этой фазы, имеющей конкретное морфологическое и биохимическое выражение. ■ Следует заметить, что сохранение альтеративной фазы воспалительной реакции не оправдывает выделения альтеративной формы воспаления, при которой сама сосудисто-мезенхимальная реакция на повреждение практически отсутствует. Поэтому необходимо согласиться с большинством современных патологов в том, что признание альтеративного воспаления, выделяемого классической патологией прошлого, противоречит сути воспалительной реакции в ее современном толковании. Повреждение и медиация — неразрывные компоненты морфогенеза воспаления, поскольку медиаторы "рождаются" в самом повреждении (альтерации). Принято выделять плазменные (циркулирующие) медиаторы, представленные прежде всего калликреин-кининовой системой, системой комплемента и системой свертывания крови, а также клеточные (локальные) медиаторы, связанные со многими клетками: лаброцитами, тромбоцитами, базофилами, ПЯЛ, макрофагами, лимфоцитами, фибробластами и др. Однако и плазменные, и клеточные медиаторы тесно взаимосвязаны и работают при воспалении как аутокаталитическая система, использующая принципы "обратной связи", "дублирования", "необходимого разнообразия" и"антагонизма". Эти принципы системы позволяют циркулирующим медиаторам обеспечить повышение сосудистой проницаемости и активацию хемотаксиса ПЯЛ для фагоцитоза, а внутрисосудистую коагуляцию в отводящих из очага воспаления сосудах — для отграничения возбудителя и самого очага воспаления (барьерная функция очага воспаления). При этом основные этапы сосудистой реакции — повышение проницаемости, активация хемотаксиса ПЯЛ и фактора Хагемана — дублируются несколькими медиаторами. Те же принципы системы в аутокаталитической реакции клеточных медиаторов обеспечивают не только повышение сосудистой проницаемости, фагоцитоз и вторичную деструкцию, но и включение иммунного ответа для элиминации повреждающего агента и продуктов повреждения и, наконец, репарацию ткани путем пролиферации и дифференцировки клеток в очаге воспаления. Наиболее ярко принцип дублирования выражен среди клеток — носителей вазоактивных веществ — лаброцитов, базофи-лов, тромбоцитов, а антагонистические начала — между этими клетками и эозинофильными лейкоцитами: медиаторы лаброцитов и базофилов стимулируют хемотаксис эозинофилов, последние же способны инактивировать эти медиаторы и фагоцитировать гранулы лаброцитов (схема 17). Среди клеток, несущих медиаторы сосудистой проницаемости, возникает "антагонистическое равновесие", определяющее своеобразие морфологии сосудистой фазы воспаления, особенно при аллергических реакциях. Клеточные медиаторы — лейкокины, монокины (интерлей-кин-1), лимфокины (интерлейкин-2) и фиброкины — являются локальными регуляторами кооперации клеток на "поле" воспаления — ПЯЛ, макрофага, лимфоцита и фибробласта [Серов В.В., Шехтер А.Б., 1981]. Другими словами, клеточные медиаторы определяют последовательность и долю участия в воспалении фагоцитарной и иммунной систем, с одной стороны, и системы соединительной ткани — с другой. "Дирижером" ансамбля клеточных медиаторов следует считать монокины макрофагов (схема 18). Макрофаги,
поддерживаемые медиаторной ауторегуляцией, способны управлять с помощью монокинов дифференцировкой гранулоцитов и моноцитов из стволовой клетки, пролиферацией этих клеток, т.е. являются регуляторами фагоцитоза. Макрофаги не только влияют на функциональную активность Т- и В-лимфоцитов, принимают участие в их кооперации, но и секретируют 6 первых компонентов комплемента, т.е. являются посредниками привлечения иммунной системы в воспалительную реакцию. Макрофаги индуцируют рост фибробластов и синтез коллагена, т.е. являются стимуляторами завершающей фазы репаративной реакции при воспалении. Вместе с тем сами макрофаги испытывают регулярное влияние лимфокинов и фиброкинов, т.е. теснейшим образом связаны в локальной клеточной регуляции с лимфоцитом и фиб-робластом [Серов В.В., Шехтер А.Б., 1981; Маянский А.Н., Ма-янский Д.Н., 1983].
Клеточная рецепция играет огромную роль в локальной клеточной регуляции при воспалении. С нею связаны межклеточное взаимодействие и привлечение в очаг воспаления компонентов иммунных реакций, так как у всех эффекторных клеток воспаления обнаружены Fc-рецепторы иммуноглобулинов и С-рецепторы комплемента. Становятся понятными неразрывная связь и неравнозначное во времени сопряжение фагоцитарной системы, иммунной системы и системы соединительной ткани в реализации конечной цели воспалительной реакции (схема 19).
Варианты этого сопряжения, зависящие от особенностей как повреждающего агента, так и организма, реагирующего на повреждение, должны, вероятнее всего, определять развитие той или иной формы воспаления. Так, гнойное воспаление (вид экссу-дативного воспаления) отражает, вероятно, особую форму сопряжения функционально несостоятельной системы ПЯЛ с макрофагами. При этом макрофаги, усиленно фагоцитирующие распадающиеся ПЯЛ, становятся устойчивыми к возбудителю. В.Е.Пигаревский (1978), изучающий это особое взаимоотношение двух систем фагоцитоза, называет его резорбтивной клеточной резистенцией. Как видно, оно отражает вторичную несостоятельность фагоцитарной функции макрофагов при первичной несостоятельности фагоцитоза ПЯЛ. Первичная и избирательная несостоятельность системы мо-ноцитарных фагоцитов, разобщение ее с системой ПЯЛ лежат в основе гранулематозного воспаления (вид продуктивного воспаления). Фагоцитарная недостаточность макрофагов определяет образование из них эпителиоидных и гигантских клеток, теряющих фагоцитарные функции. Фагоцитоз подменяется отграниче- нием, персистенцией возбудителя. Незавершенный фагоцитоз делает незавершенной и несовершенной саму воспалительную реакцию. Она становится выражением реакции гиперчувствительности замедленного типа (ГЗТ). Очевидно также, что наследственные дефекты каждой из систем защиты, как и системы самой соединительной ткани, делают дефектной и воспалительную реакцию как по форме ее проявления и течению, так и по возможности реализации конечной цели. Достаточно вспомнить наследственную недостаточность бактерицидных систем ПЯЛ и моноцитов, наиболее ярко представленную при хронической гранулематозной болезни детей, наследственные и врожденные иммунные дефициты и фатальность развивающейся при них гнойной инфекции, врожденную несостоятельность соединительной ткани и упорство хронического воспаления. Нельзя не сказать и о наследственных дефицитах системы комплемента, особенно СЗ- и С5-компонснтов ее. Эти дефициты проявляются либо рецидивирующей гнойной инфекцией, либо волчаночноподобным синдромом. При воспалении, особенно вызываемом различными агентами, пошлются как циркулирующие в крови, так и локальные гетсрологичные иммунные комплексы, при хроническом течении воспалении они могут быть и аутологичными. Так, при воспалении возникают иммунокомплексные реакции — наиболее частые среди реакций гиперчувствительности немедленного типа (ГНТ).
Поиск по сайту: |